
| T细胞 | |
|---|---|
 人类T细胞的扫描电子显微镜图
| |
| |
| 基本信息 | |
| 系統 | 免疫系统 |
| 标识字符 | |
| 拉丁文 | lymphocytus T |
| MeSH | D013601 |
| TH | H2.00.04.1.02007 |
| FMA | FMA:62870 |
| 《显微解剖学术语》 | |
T细胞(英語:T cell、T lymphocyte)是淋巴细胞的一种,在免疫反應中扮演着重要的角色。T是胸腺(thymus)的英文缩写。T细胞在骨髓被製造出來之後,在胸腺内進行「新兵訓練」分化成熟為不同亚型的效应T細胞,成熟后就移居于周围淋巴组织中開始工作。
T细胞膜表面分子有两大类:T细胞受体(TCR)和分化群分子(CD molecules),其与T细胞的功能相关,也是T细胞的表面标志(cell-surface marker),可以用以分离、鉴定不同亚群的T细胞。
发育
起源
所有的T细胞都来源于造血干细胞(HSC),造血干细胞之后会分化为多能祖细胞(MPP),多能祖细胞又会分化为共同淋巴祖细胞(CLP),CLP接下来只有三种分化路径,即T细胞、B细胞和NK细胞。 那些分化为T细胞的CLP将会随着血流到达胸腺,并成为早期胸腺祖细胞(ETP),现在这些细胞既不表达CD4也不表达CD8。这些细胞将经过一轮分裂,进入DN1阶段。
TCR-β选择
在DN2阶段(CD44+CD25+),细胞上调RAG1/2并重排TCR(T细胞受体)-β基因座,V-D-J序列和恒定区序列,目的是产生一个有功能的TCR-β链。当细胞经过DN3阶段(CD44-CD25+)时,细胞将会和TCRβ一起表达一个未经重排的α-链(pre-Tα),如果重排后的β-链可以和pre-Tα形成二聚体,细胞将产生信号停止β-链的重排。虽然这个信号需要pre-TCR在细胞膜上表达,不过它与pre-TCR和配体的结合无关。如果pre-TCR形成了,细胞会下调CD25并进入DN4阶段(CD25-CD44-)。这些细胞将继续分裂并重排TCRα的基因座。
阳性选择
双阳性(CD4+/CD8+)的T细胞会向胸腺皮层深处迁移,并会接触到胸腺皮层上皮细胞表面的“自体抗原”(self-antigens)。这些自体抗原结合在胸腺上皮细胞表面的MHC分子上,只有与胸腺细胞的MHC分子表现出足够强的结合力的T细胞,才能接收到必要的“存活信号”,而无法接收到足够“存活信号”的T细胞将会凋亡。在这个持续几天的阳性选择过程中,大部分的T细胞都会死去。
一个T细胞的命运就在阳性选择的过程中被决定。在双阳性(CD4+/CD8+)T细胞中,能够与MHC Ⅱ类分子结合得较好的将成为CD4+细胞,而和MHC Ⅰ类分子有更高亲和力的将成为CD8+细胞。将成为CD4+细胞的细胞将会逐渐下调自己的CD8,最终成为单阳性的CD4+细胞。
阴性选择
在阳性选择中存活下来的T细胞将会向胸腺皮质边缘和髓质区迁移;在髓质区,它们又会接触到胸腺髓质上皮细胞(mTECs)表面的自体抗原。mTECs会在它们的MHC Ⅰ类分子上呈递来自全身各个组织的自体抗原。一些mTECs被胸腺树状细胞吞噬,它们的自体抗原就会呈递在树状细胞的MHC Ⅱ类分子上(经过了阳性选择的CD4+细胞只能识别MHC Ⅱ类分子)。在这里,与自体抗原表现出过强的亲和力的T细胞会接收到凋亡信号并凋亡(在这些细胞中也有一部分会成为调节T细胞),存活下来的细胞就作为成熟的初级T细胞离开胸腺。这一过程是中枢免疫耐受的重要组成部分,其意义在于筛选掉可能对自体抗原产生反应的T细胞,从而避免自体免疫疾病的发生。
胸腺输出
经过阳性选择和阴性选择,最初到达胸腺的T细胞中有98%死亡,存活下来的2%成为了具有成熟免疫功能的T细胞。胸腺产生成熟T细胞的数量大致随着个体衰老而减少,在中年人的体内,胸腺的大小平均每年缩小3%。所以,对中老年人而言,外周T细胞的增殖和再生对于免疫系统的意义更大。
分类
T细胞根据功能的差异被分为几个亚型。虽然在胸腺中就分化出了CD4+和CD8+两者,但是在外周T细胞还会发生进一步的分化。
常规适应性T细胞
辅助性CD4+ T细胞
辅助性T细胞(TH细胞)对其他淋巴细胞的活动起辅助作用,包括B细胞向浆细胞和记忆B细胞的发育,以及细胞毒性T细胞和巨噬细胞的激活。它们也被称为CD4+ T细胞,因为它们的细胞表面有CD4蛋白的表达。辅助T细胞在遇到抗原呈递细胞(APC)表面MHC-II分子结合的外部抗原时被激活,一旦被激活就会快速分裂并开始分泌调节免疫反应的细胞因子。辅助T细胞在受到不同细胞因子刺激的情况下,也会进一步分化成不同亚型的辅助T细胞。它们是已知的HIV病毒的目標细胞,在艾滋病发病时会急剧减少。滤泡辅助性T细胞是参与体液应答的CD4阳性辅助性T细胞的一个亚群,属一类专门为B细胞提供帮助的CD4+ T细胞。
調節性CD4+ T细胞
[調節性T细胞](Treg细胞)旧称抑制性T细胞,对于免疫耐受至关重要。它们的主要工作就是及时有效的结束免疫反应,以及抑制那些从阴性筛选中逃逸的自体免疫T细胞,防止免疫反应对抑制性机体自身造成过度损害。
調節性T细胞既可以在胸腺中发育分化完成,称为胸腺调节T细胞;也可以在外周组织受免疫反应诱导分化,称为外周调节T细胞。两者都表达FOXP3作为其细胞表面标志物,FOXP3基因的突变会影响调节T细胞的发育,并诱发致命的自体免疫病IPEX。
其他几类不表达FOXP3基因的T细胞具有免疫抑制作用,例如Tr1细胞和Th3细胞。Tr1与IL-10相关,Th3与TGF-beta相关。最近,Th17细胞也被列入此类免疫抑制细胞之中。
细胞毒性CD8+ T细胞
细胞毒性T细胞(CTLs, killer T cells)负责杀伤被病毒感染的细胞和癌细胞,在对器官移植的免疫排斥中也有参与。其特点在于细胞表面的CD8蛋白质。它通过识别所有有核细胞表面的MHC-I分子上的短肽抗原,来分辨正常细胞和应杀伤的异常细胞。细胞毒性T细胞还可分泌重要的细胞因子IL-2和IFNγ,来影响其他免疫细胞的功能,特别是巨噬细胞和NK细胞。
记忆性T细胞
还未结合过外部抗原的初级T细胞,一旦结合了抗原呈递细胞表面MHC分子所包裹的外部抗原,就会开始增殖分化为效应T细胞和记忆T细胞(其他信号适当的共刺激对这一过程也是必要的)。曾经,人们认为记忆T细胞只分为中央记忆T细胞和效应记忆T细胞。但是之后,新的记忆T细胞种类不断被发现,例如组织驻留记忆T细胞 (Trm)等等。记忆T细胞的共同特点在于其寿命较长(可長達數十年),而且在识别到特定抗原时可以快速分裂为大量的效应T细胞。通过这样的方式,记忆T细胞就为人体的免疫系统保存了对之前感染过病原体的“记忆”。记忆T既可以是CD4+也可以是CD8+,一般会表达CD45RO。
类固有T细胞
自然杀伤T细胞
自然杀伤T细胞(NKT细胞),请不要把它和固有免疫中的自然杀伤细胞(NK细胞)混淆。 与一般T细胞识别MHC分子上的肽链抗原不同,NKT识别的是CD1d分子上结合的糖蛋白抗原。被激活后,它们可以执行类似辅助T细胞和细胞毒性T细胞的功能,即释放细胞因子和细胞毒素。有证据表明,它们能够识别并杀灭某些肿瘤细胞和被疱疹病毒感染的细胞。
粘膜相关的不变T细胞
粘膜相关的不变T细胞(MAIT)具有固有免疫效应细胞的特质。在人体内,MAIT细胞分布于血液、肝脏、肺部、黏膜,具有抵御微生物感染的能力。MHC-I的类似物,MR1,可以向MAIT细胞呈递细菌产生的代谢物抗原。接受了MR1呈递的外部抗原后,MAIT细胞可以释放促炎症细胞因子并裂解被细菌感染的细胞。MAIT也可以通过不依靠MR1的信号通路激活。除了表现出类似固有免疫的功能外,MAIT细胞也辅助获得性免疫反应,甚至表现出类似记忆细胞的特征。此外,MAIT也被认为在自体免疫病中发挥作用,如多发性硬化、风湿和炎症性肠病,虽然决定性的证据还有待发现。
γδ T细胞
γδ T细胞代表了T细胞中一小部分不表达αβ-TCR而表达γδ-TCR的类型,在小鼠和人体内仅占全部T细胞的2%;在兔子、绵羊和鸡体内,γδ T细胞占全部T细胞的比例则可能高达60%。它们主要分布在肠道粘膜,作为一类上皮内淋巴细胞。关于其抗原识别的细节我们仍知之甚少,不过似乎γδ T细胞并不受限于MHC分子的呈递。特别的是,γδ T细胞能够对一类磷酸类抗原做出快速的反应,而这类抗原物质在各种细胞(细菌、植物、癌细胞等)中都有发现。
活化
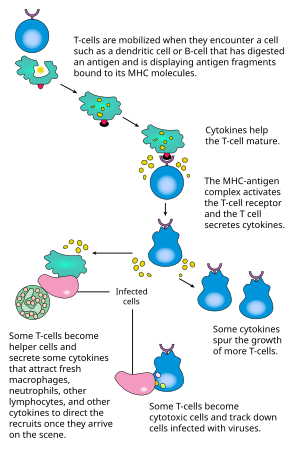
CD4+ T细胞的激活需要T细胞上的TCR和共受体(CD28或ICOS),抗原呈递细胞上的MHCII和共激活分子两对分子的分别,同时结合。仅其中一对的结合,无法产生有效的T细胞激活。理想的CD8+ T细胞激活则依赖于CD4+ T细胞的信号转导。CD4+细胞可以在初级CD8 T细胞的初次免疫应答中给予帮助,并且在急性感染的后期维持CD8+ 记忆T细胞的活性。所以,CD4+ T的激活对于CD8+ T细胞的活动是有利的。
相比于MHC分子上的抗原,抗原呈递细胞的共激活分子一般是由病原体的副产物、热休克蛋白或者坏死的细胞碎片诱导表达的。共刺激机制被认为可以避免自体免疫的发生,因为即使T细胞错误地结合了自体抗原,也可能因为没有受到合适的共刺激而无法正常活化。一旦T细胞被正确地活化,它的细胞表面蛋白表达就会发生巨大的改变,活化T细胞的标志蛋白包括CD69,CD71,CD25 (也是调节T细胞的标志)和HLA-DR (人类T细胞的特异标志)。CTLA-4在活化T细胞表面的上调,对共激活受体有竞争性抑制作用,可以避免活化T细胞的过度活化。活化T细胞的表面糖基化情况也有改变。
T细胞受体(TCR)是由几种蛋白质组合成的复合体。TCR的两个主要组分是由两个独立基因分别编码的TCRα和TCRβ,其他的组分包括CD3家族的蛋白:CD3εγ和CD3εδ的异二聚体,以及最重要的CD3ζ同二聚体。CD3ζ同二聚体上共有6个ITAM基序,可被磷酸化并启动一系列级联反应,导致TCR复合体的聚集。
虽然在绝大部分情况下T细胞活化都依赖于TCR对抗原的识别,其他的活化途径也有被发现,例如细胞毒性T细胞可以被其他CD8 T细胞识别并导致自身的极化。T细胞活化的过程也受到活性氧类物质的影响。
抗原识别
T细胞的主要特点就是能够分辨正常细胞和异常细胞的能力。不论是正常细胞还是异常细胞,都会表达大量的MHC-抗原多肽复合体(pMHC)。虽然T细胞与正常细胞的pMHC有一定结合力,但是T细胞并不会被激活;但即使异常细胞的pMHC与正常细胞只有细微的差别,也能够刺激T细胞发生免疫反应。这样对不同抗原完全不同的反应特征称为T细胞的抗原识别,关于这一机理实现的具体细节如今仍然没有定论。
临床意义
缺陷
T细胞缺陷可能意味着T细胞数量的减少或者T细胞功能的缺失。完全的T细胞缺陷可能来自于一些遗传因素,例如严重复合型免疫缺乏症(SCID)、欧门氏症候群或软骨毛发发育不全。[46]部分的T细胞缺陷可能是由于获得性免疫缺陷综合征(AIDS)、遗传性的迪乔治综合征(DGS)、染色体断裂综合征(CBSs),或者B细胞和T细胞的复合缺陷,例如毛细血管扩张性运动失调 (AT) 和欧德里综合征(Wiskott–Aldrich syndrome)。
T细胞缺陷患者面临的主要风险,主要是一些细胞内病原体,例如单纯疱疹病毒、分枝杆菌和李斯特菌。同时,真菌感染在T细胞缺陷患者身上往往也很常见且严重。
癌症
T细胞癌变诱发的肿瘤称为T细胞淋巴瘤,在非霍奇金淋巴瘤中约占10%。
耗竭
T细胞耗竭是一种T细胞功能失常的状态,其表现为进展性的功能丧失、基因表达谱的变化、和抑制性细胞因子的持续分泌。T细胞耗竭可能发生于慢性感染、败血症、癌症的进程中。耗竭的T细胞即使再次暴露于抗原刺激之中也无法恢复正常功能。
慢性感染和败血症
T细胞耗竭的直接原因包括持续的抗原刺激、以及CD4细胞的缺失。长时间的抗原暴露和高病毒负载可以加重T细胞耗竭的程度。2-4周的持续抗原暴露就可导致T细胞耗竭。另一个可以导致T细胞耗竭的因素是包括PD-1在内的一系列抑制性受体。细胞因子IL-10或TGF-β也可以导致耗竭。调节T细胞因为可以分泌IL-10和TGF-β,也与T细胞耗竭相关。在阻断PD-1受体并减少调节T细胞数量后,T细胞耗竭的情况可以得到反转。[58]在败血症中,抑制性的细胞因子风暴也会造成T细胞耗竭。现在已有致力于通过阻断抑制性受体的方式来治疗败血症的疗法研究。
器官移植
与感染时的情况类似,器官移植带来的持续异种抗原暴露也会造成T细胞耗竭。肾移植后,T细胞应答能力会随时间减弱。这些数据说明T细胞耗竭导致的CD8+ T细胞数量减少可能是器官移植耐受中的重要一环。已有几项研究证明了慢性感染对器官移植后的免疫耐受和长期生存有利,而T细胞耗竭起着一定介导的作用。虽然已有T细胞耗竭对器官移植有利的证据,但是T细胞耗竭同时带来的感染和癌变风险依然不能忽视。
癌症
在癌症进程中,T细胞耗竭显然对癌组织的存活有利。已有研究证明癌细胞和一些癌症相关细胞可以主动地诱导T细胞耗竭的发生。在白血病中,T细胞耗竭也与其复发相关。一些研究甚至提出可以基于T细胞抑制性受体PD-1的表达状态来预测白血病复发的情况。由于免疫抑制性受体与T细胞耗竭以及癌症之间的关系,近年来有大量的研究和临床试验致力于通过阻断免疫抑制性受体来治疗癌症,其中有一些已经被认定有效并投入临床使用。
参见
外部連結
- Immunobiology, 5th Edition(页面存档备份,存于互联网档案馆)
- niaid.nih.gov – The Immune System
- T-cell Group – Cardiff University
- (Successful!) Treatment of Metastatic Melanoma with Autologous CD4+ T Cells against NY-ESO-1(页面存档备份,存于互联网档案馆).
- The Center for Modeling Immunity to Enteric Pathogens (MIEP)(页面存档备份,存于互联网档案馆)
- Anthony J. Davies. The tale of T cells. Immunology Today: 137–140. [2018-04-02]. doi:10.1016/0167-5699(93)90216-8. (原始内容存档于2018-06-29).
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||