
| 遗传学 |
|---|
 |
| 重要概念 |
| 历史及分支 |
| 研究 |
| 個人化醫療 |
| 精準醫學 |
| 生物學的一部分 |
| 生物化学 |
|---|
 |
| 关键部分 |
| 歷史和主題 |
| 詞彙 |
| 生物化學主题 |
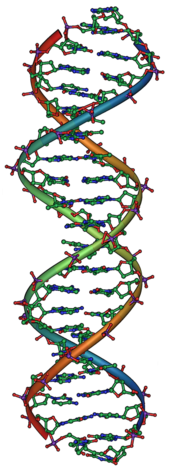
遗传学是研究生物体的遗传和变异的科学,是生物学的一个重要分支。史前时期,人们就已经利用生物体的遗传特性通过选择育种来提高谷物和牲畜的产量。而现代遗传学,其目的是寻求了解遗传的整个过程的机制,则是开始于19世纪中期孟德尔的研究工作。虽然孟德尔并不知道遗传的物理基础,但他观察到了生物体的遗传特性,某些遗传单位遵守简单的统计学规律,这些遗传单位现在被称为基因。
基因位于DNA上,而DNA是由四类不同的核苷酸组成的链状分子,DNA上的核苷酸序列就是生物体的遗传信息。天然DNA以双链形式存在,两条链上的核苷酸互补,而每一条链都能够作为模板来合成新的互补链。这就是生成可以被遗传的基因的复制方式。
基因上的核苷酸序列可以被细胞翻译以合成蛋白质,蛋白质上的氨基酸序列就对应着基因上的核苷酸序列。这种对应性被称为遗传密码。蛋白质的氨基酸序列决定了它如何折叠成为一个三维结构,而蛋白质结构则与它所发挥的功能密不可分。蛋白质执行细胞中几乎所有的生物学进程来维持细胞的生存。DNA上的一个基因的改变可以改变其编码的蛋白质的氨基酸,并可能改变此蛋白质的结构和功能,进而对细胞甚至整个生物体造成巨大的影响。
虽然遗传学在决定生物体外形和行为的过程中扮演着重要的角色,但此过程是遗传学和生物体所经历的环境共同作用的结果。 例如,虽然基因能够在一定程度上决定一个人的体重,人在孩童时期的所经历的营养和健康状况也对他的体重有重大影响。
历史

虽然遗传科学开始于格里哥·孟德尔在19世纪中期的工作(包括实验和理论),但其他一些关于遗传的理论研究则早于孟德尔。在孟德尔时期,一种比较流行的理论——“混合遗传”(blending inheritance)提出:个体的遗传特征是来自于其父母的特征的混合平均值。孟德尔的工作则否定了这一理论,他的结果显示遗传特征是由不同基因综合表现的结果而不是连续的混合。当时的另一种得到人们支持的理论——“用进废退说”(又称为“获得性遗传”)提出生物经常使用的器官逐渐发达,不使用的器官逐渐退化,并且这种后天获得的性状是可以遗传的。这一由拉马克所提出的理论后来被证明是错误的,因为通常个体的经历并不影响它们的基因,也就不会遗传给下一代。(注:但近年来对于表观遗传现象的研究发现,由于不同的经历而引起的不同的性状在一些情况下是可以被遗传的,虽然这些发现并不能证明拉马克的理论。)
古典遗传学

现代遗传学的奠基者是格里哥·孟德尔,一个奥地利修道士和科学家。他致力于研究植物的遗传现象。1865年,他的论文《Versuche über Pflanzenhybriden》(《植物杂交实验》)发表在布尔诺的自然研究学会(Naturforschender Verein)上。在论文中,他展示了豌豆在杂交实验中所表现的遗传规律并以数学关系加以描述。 虽然这种遗传规律只能在表现类型数量很少的情况下才能被观察到,但是孟德尔的工作显示遗传是颗粒性的(颗粒遗传,与混合遗传相对,所谓的颗粒相当于现在为人们所知的基因),而不是混合性或获得性的,并且许多性状的遗传规律可以通过简单的规则和比率来解释。
孟德尔工作的重要性并没有得到广泛的理解,直到他逝世之后的1890年代,当雨果·德弗里斯遇到相同的情况时才重新发掘出他的研究结果。作为孟德尔理论的支持者,威廉·贝特森(William Bateson)提出了genetics(“遗传学”)这一名词。 1906年,伦敦召开的第三次国际植物杂交大会上,在贝特森的提议下,genetics这一单词得以广泛使用于描述关于遗传的研究。
在重新评价孟德尔的工作后,科学家们试图确定细胞中的哪一种分子是遗传物质。1910年,基于对果蝇的性连锁白眼突变的观察结果,托马斯·亨特·摩尔根提出基因位于染色体上。 1913年,他的学生阿尔弗雷德·斯特蒂文特利用遗传连锁的现象显示了基因是在染色体上呈线性排列的。
分子遗传学
虽然摩尔根等人的工作使得人们认识到基因是位于染色体上,但染色体是由蛋白质和DNA共同组成的,研究者们依然不知道哪一种物质才是遗传物质。1928年,弗雷德里克·格里菲斯发现了转化现象(参见格里菲斯實驗):死亡的细菌可以将遗传物质“转化”到其他依然活着的细菌内。16年后的1944年,奥斯瓦尔德·埃弗里、科林·马克聊德(Colin McLeod)和马克林·马克卡提(Maclyn McCarty)鉴定出进行转化的物质是DNA。 1952年,赫希-蔡斯實驗再次显示是DNA(而不是蛋白质)才是感染细菌的病毒的遗传物质,从而进一步证明了DNA是遗传信息的携带者。
1953年,利用羅莎琳·富蘭克林对DNA进行的X射线晶体学的研究成果,詹姆斯·杜威·沃森和弗朗西斯·克里克成功解析了DNA的双螺旋结构。 他们所提出的双螺旋模型包含有两条DNA链,链之间通过核苷酸上的碱基配对,从而形成一个類似于旋转梯子状的结构。 DNA结构显示了遗传信息存在于每条DNA链的核苷酸序列中。这一结构也提示了一种简单的DNA复制方法:两条配对的DNA链分开后,新的配对链可以根据旧链上的序列来搭建。
虽然DNA结构显示了遗传的进行方式,但人们依然不知道DNA是如何影响细胞行为的。随后的多年时间中,科学家们试图了解DNA是如何控制蛋白质的制造过程。科学家发现细胞利用DNA作为模板来生成配对的信使RNA(RNA是一种类似于DNA的分子)。信使RNA上的核苷酸信息就被用于生产蛋白质上的氨基酸序列;这种由核苷酸序列到氨基酸序列的翻译是根据遗传密码的规则进行的。
随着遗传的分子机制的揭示,大量的研究成果不断涌现。其中,一个重要的发展是弗雷德里克·桑格于1977年提出的链终止DNA测序法,这一方法使得科学家们可以阅读DNA分子上的核苷酸序列。 1983年,凯利·穆利斯发展出了聚合酶链锁反应技术,从而为从混合物中分离和扩增特定的DNA提供了一个快捷而灵敏的方法。 经过人类基因组计划和同时的竞争者塞雷拉基因組(一个私人赞助的基因组计划)的努力,人类基因组的测序在2003年得以基本完成。
遗传的特点
不连续遗传和孟德尔定律
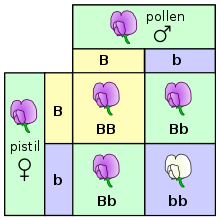
在最基本的水平上,生物体中的遗传表现为离散性状,即基因型。 这种特点是由孟德尔首次观察到,他研究了豌豆中遗传性状的分离现象。 在研究花色的实验中,孟德尔观察到豌豆花的颜色只有两种:紫色和白色,却没有任何一朵显示出两种颜色的中间色。这些来自于同一基因却不同且离散的版本被称为等位基因。
在豌豆的例子中,每一颗豌豆都含有一个基因中的两个等位基因,并且子代可以从父母分别继承其中的一个等位基因。 许多生物,包括人类,都有这样的遗传规律。具有相同的两个等位基因的生物体被称为纯合体,而具有不同等位基因的生物体则被称为杂合体。
一个给定的生物体的等位基因的组合形式就是该生物体的基因型,而对于这种组合所表现出来的性状就是该生物体的表现型。当生物体是杂合体时,常常有一个等位基因是显性基因,显性基因决定了生物体的表现型,而另一个基因就被称为隐性基因,其性状在显性基因存在时不会被表现出来。有一些等位基因没有完全的显性,即“非完全显性”,其表现为一种中间状态的表现型,或者两个等位基因无显隐性之分可以同时表现出对应性状。
当一对生物体繁殖后代时,它们的下一代随机地继承父母的两个等位基因中的一个。这些对于离散遗传和等位基因分离的观察结果被总结为孟德尔第一定律(分离定律)。
注释和图解

遗传学家利用注释和图解来描述遗传。一个基因可以用一个或几个字母来表示,并且用大写字母表示显性基因,小写字母表示隐性基因。“+”常常被用于标识一个基因的正常非突变的等位基因。
在杂交实验中(特别是在讨论孟德尔定律时),父母代被标示为“P”代,其下一代标示为“F1”(第一代)。F1代的子代就被称为“F2”(第二代)。可以用于预测杂交结果的常用图解是旁氏表(Punnett square,又称为“棋盘法”)。
在研究人类遗传疾病时,遗传学家常常利用系谱图来展示遗传性状。 这些图表将一个性状的遗传关系以家族谱(family tree)的形式表现出来。
多基因的相互作用
生物体具有成千上万个基因,并且在有性繁殖的生物中,这些基因的分类是互相独立的。这就意味着对应豌豆的黄色或绿色的色彩等位基因的遗传与紫色或白色的花色等位基因的遗传是不相关的。这种现象被称为孟德尔第二定律(又称为“独立分配定律”),即来自父母的不同基因的等位基因被随机抽取来组成具有多种组合结果的子代。有一些基因不是独立归类的,这也就解释了遗传中的遗传连锁现象(具体内容见后)。
不同的基因常常能够通过某种方式来影响同一种性状,例如春锦龙花(Collinsia verna)具有一种决定花色为蓝色或洋紅色的基因,以及一种能够决定花是否有颜色(即白色或有色)的基因。当一株春锦龙花含有两个白色等位基因(决定花色的第二种基因的两个等位基因),则无论第一种基因所带的颜色基因的等位基因为何,它的花色都为白色。这种基因之间的作用关系被称为上位性或异位显性(epistasis),即第二种基因位于第一种基因的上位。
许多性状没有明显的可区分的特点(例如不同的花色),而表现为连续性的特点(如人类的身高和肤色)。这些复杂的性状是来自于许多基因共同作用的结果。 这些基因的影响作用在不同程度上是由一个生物体所经历的环境所介导的。生物体的一个基因对于一种复杂性状产生的影响的程度被称为遗传力。 对一种复杂性状的遗传力的测量是相对的:环境的变化性越大,环境对于性状变化的影响力也就增强,而基因对于性状变化的影响力也就越小(表现为遗传力降低)。例如,作为一种复杂性状,美国人身高的遗传力为89%;而在尼日利亚,由于人们所获得的食物和医疗保健的差异性较大(即较大的环境变化性),其身高的遗传力仅为62%。
遗传的分子基础
DNA和染色体


基因的分子基础是脱氧核糖核酸(DNA)。DNA由核苷酸相互连接而形成的链分子,其中的核苷酸有四类:腺苷酸(A)、胞嘧啶(C)、鸟苷酸(G)和胸腺嘧啶(T)。遗传信息就储存在这些核苷酸序列中,而基因则以连续的核苷酸序列存在于DNA链中。病毒是唯一的例外,有一些病毒利用核糖核酸(RNA)分子来代替DNA作为它们的遗传物质。
DNA通常以双链分子的形式存在,并卷曲形成双螺旋结构。DNA中的每一个核苷酸都有自己的配对核苷酸在相反链(对应另一条链)上,其配对规则为:A与T配对,C与G配对。因此,双链中的每一条链都包含了所有必要的遗传信息。这种DNA结构就是遗传的物理基础:DNA复制通过将互补配对的双链分开并利用每条链作为模板来合成新的互补链,从而达到复制遗传信息的目的。
不同基因沿着DNA链线性排列形成了染色体。在细菌中,每一个细胞都有一个单一的环状染色体;而真核生物(包括动物和植物)则具有多个线形染色体。这些染色体中的DNA链常常会非常长;例如,人类最长的染色体的长度大约为247百万个碱基对。 染色体DNA上结合有能够组织和压缩DNA并控制DNA可接触性的结构蛋白,从而形成染色质;在真核生物中,染色质通常是以核小体为单位组成,每一个核小体由DNA环绕一个组蛋白核心而形成。 一个生物体中的全套遗传物质(通常包括所有染色体中DNA的序列)被称为基因组。
仅含有一套染色体的生物被称为单倍体生物;大多数的动物和许多植物为双倍体生物,它们含有两套染色体(姐妹染色体),即含有每个基因的两个拷贝。 一个基因的两个等位基因分别位于姐妹染色体上的等同的基因座,每一个等位基因遗传自不同亲本。

性染色体是双倍体生物中染色体的一个例外,它是许多动物中的一种特异化的染色体,决定了一个生物体的性别。 在人类和其他一些哺乳动物中,性染色体分为X和Y两类。Y染色体只含有很少量的基因,能够触发雄性特征的发育;而X染色体与其他染色体类似,也含有大量与性别决定无关的基因。雌性具有两个X染色体,而雄性具有一个Y染色体和一个X染色体。这种X染色体拷贝数的差别是性连锁的遗传病具有特殊遗传规律的原因。
繁殖
当细胞分裂时,它们的基因组被复制产生两份拷贝,每个子细胞继承其中的一份。这一过程被称为有丝分裂,它是繁殖的最简单形式,也是无性繁殖的基础。无性繁殖也能够发生在多细胞生物体中,子代从单一亲本处继承其基因组,即子代与亲本具有等同的基因组。这种子代与亲本在遗传上等同的现象被称为克隆。
真核生物常常利用有性繁殖来产生后代,其后代含有分别遗传自不同亲本的混合的遗传物质。有性繁殖的过程是一个介于基因组单拷贝(单倍体)和双拷贝(双倍体)之间的一个转换过程。 双倍体生物通过不复制DNA的分裂来形成单倍体,所生成的单倍体子细胞含有每对姐妹染色体中的任意一个。两个单倍体细胞融合并将各自的遗传物质组合在一起来重新生成一个含配对染色体的双倍体细胞。多数动物和许多植物在它们的生命周期的多数时间内是双倍体,只有生殖细胞为单倍体形式。
虽然细菌没有单倍体/双倍体的有性繁殖方式,它们也有许多获得新的遗传信息的手段。一些细菌能够发生接合,将一小段环状DNA传递到另一个细菌细胞内。 细菌还能够从环境中摄入DNA片断,并将之整合到自己的基因组中,这种现象被称为转化。 这些进程导致了基因的水平转移,即无亲缘关系的生物体之间进行遗传信息的传输。
重组与连接

染色体的双倍体使得位于不同染色体的基因在有性繁殖期间能够独立地分配(参见独立分配定律),并通过重组形成新的基因组合。在同一条染色体上的基因理论上不会发生重组,但通过染色体互换则可以达到。在互换过程中,染色体交换DNA片段,有效地将染色体之间的等位基因重新分配。 染色体互换通常发生在减数分裂期间(旨在生成单倍体细胞的一系列的细胞分裂过程)。
染色体上两个给定位点之间发生染色体互换的可能性与这两个位点之间的距离相关。对一个任意长的距离,由于互换的可能性足够高,使得相隔该距离的两个基因的遗传无相关性。对于两个接近的基因,由于互换的可能性较小,则基因之间会发生遗传连锁,即这两个基因的等位基因趋向于被一起遗传。一系列基因之间的连锁数量可以被组合在一起构成一个线性的连锁图谱来描述染色体上基因的排列顺序。
基因表达
遗传密码

基因通常是通过生成所编码的蛋白质(执行细胞中大多数功能的复杂的生物大分子)来表现它们的功能性影响。蛋白质是由氨基酸所组成的线性链,而基因的DNA序列(通过RNA作为信息的中间载体)被用于产生特定的蛋白质的氨基酸序列。这一过程的第一步是由基因的DNA序列来生成一个序列互补的RNA分子,即基因的转录。
通过转录产生的RNA分子(信使RNA)被用于生产相应的氨基酸序列,这一转换过程被称为翻译。核酸序列中的每一组三个核苷酸组成一个密码子,可以被翻译为20种出现于蛋白质中的氨基酸中的一个,这种对应性被称为遗传密码。 这种信息的传递是单一方向性的,即信息只能从核苷酸序列传递到氨基酸序列,而不能从氨基酸序列传递回核苷酸序列,这一现象被弗朗西斯·克里克称为分子生物学中心法则。
特定的氨基酸序列决定了对应蛋白质的独特的三维结构,而蛋白质结构则与它们的功能紧密相连。 一些蛋白质是简单的结构分子,如形成纤维的胶原蛋白。蛋白质可以与其他蛋白质或小分子结合;例如,作为酶的蛋白质通过与底物分子结合来执行催化其化学反应的功能。蛋白质结构是动态的;例如,血红蛋白在哺乳动物血液中捕捉、运输和释放氧气分子的过程中能够发生微小的结构变化。
基因序列上的单个核苷酸变化(密码子改变)可能会导致所编码蛋白质的氨基酸序列相应改变。由于蛋白质结构是由其氨基酸序列所决定的,一个氨基酸的变化就有可能通过使结构失去稳定性或改变蛋白质表面而影响与该蛋白质其他蛋白质和分子的相互作用,而引起蛋白质性质发生剧烈的改变。例如,鐮刀型細胞貧血症是一种人类遗传性疾病,是由编码血红蛋白中的β-球蛋白亚基的基因中的一个核苷酸突变所引起的,这一突变导致一个氨基酸发生改变从而改变了血红蛋白的物理性质; 在这一疾病中,突变的血红蛋白互相结合在一起,堆积而形成纤维,从而扭曲了携带血红蛋白的红血球的形状。这些扭曲的鐮刀状细胞无法在血管中通畅地流动,容易堆积而阻塞血管或者被降解,从而引起贫血疾病。
也有一些基因被转录为RNA分子后却不被翻译成蛋白质,这些RNA分子就被称为非编码RNA。在一些例子中,这些非编码RNA分子(如核糖体RNA和转运RNA)折叠形成结构并参与部分关键性细胞功能。还有的RNA(如microRNA)还能够通过与其他RNA分子杂交结合而发挥调控作用。
先天与后天

虽然基因含有生物体所需功能的所有信息,环境依然在确定生物体最终的表现型中发挥着重要作用;这种两面性被称为“先天与后天”,也可以说,一个生物体的表现型依赖于遗传与环境的相互作用。这种相互作用的一个例子就是温度敏感型突变:蛋白质序列中的单个氨基酸突变通常不会改变该蛋白质的行为和与其他分子的相互作用关系,但却能够使该蛋白质结构变得不稳定。在一个高温环境中,分子的运动加快,分子间的碰撞也加强,这就使得这一蛋白质的结构被破坏从而无法发挥它的功能;而在一个低温环境中,蛋白质结构却可以保持稳定并能够发挥正常的功能。这类突变所引起的改变在暹罗猫毛色的变化中可以被观察到。这种猫体内一种负责生产色素的酶含有一个突变,这个突变能够导致这种酶在高温时变得不稳定并失去其功能。 因此,在猫皮肤温度较低处(如四肢、尾部、面部等)的毛色为深色,而较高处为浅色。
在人类遗传疾病苯丙酮尿症中,环境因素也具有重大的影响。 导致苯丙酮尿症的突变破坏了机体降解苯丙氨酸的能力,导致具有毒性的中间产物分子在体内堆积,从而引起严重的进行性智能发育不全和癫痫。带有苯丙酮尿症突变的病人需要遵守严格的饮食,以避免摄入含苯丙氨酸的食品,才能保持正常的和健康的生活。
基因调控
一个生物体的基因组含有数千个基因,但并不是所有的基因都需要保持激活状态。基因的表达表现为被转录为mRNA,然后再被翻译成蛋白质;而细胞中存在许多方式可以来控制基因的表达,以便使蛋白质的产生符合细胞的需求。而控制基因表达“开关”的主要调控因子之一就是转录因子;它们是一类结合在基因的起始位点上的调控蛋白,可以激活或抑制基因的转录。 例如,在大肠杆菌细菌基因组内存在着一系列合成色氨酸所需的基因。然而,当细菌细胞可以从环境中获得色氨酸时,这些基因就不被细胞所需要。色氨酸的存在直接影响了这些基因的活性,这是因为色氨酸分子会与色氨酸操纵子(一种转录因子)结合,引起操纵子结构变化,使得操纵子能够结合到合成色氨酸所需基因上。色氨酸操纵子阻断了这些基因的转录和表达,因而对色氨酸的合成进程产生了负反馈调控作用。
多细胞生物中的基因表达的差异性非常明显:虽然各类细胞都含有相同的基因组,却由于不同的基因表达而具有不同的结构和行为。多细胞生物中的所有细胞都来源于一个单一细胞,通过响应外部或细胞之间的信号而不断分化并逐渐建立不同的基因表达规律来产生不同的行为。因为没有一个单一基因能够负责多细胞生物中的各个组织的发育,因此这些规律应来自于许多细胞之间的复杂的相互作用。这些过程都要通过基因调控来完成。
真核生物体内的染色质中存在着能影响基因转录的结构特点,常常表现为DNA和染色质的修饰形式(如DNA的甲基化),而且能够稳定遗传给子细胞。 这些特点是“附加性”的,因为它们存在于DNA序列的“顶端”并且可以从一个细胞遗传给它的下一代。由于这些附加性特点,在相同培养基中生长的不同的细胞类型依然保持其不同的特性。虽然附加性特点在整个发育过程中通常是动态的,但是有一些,例如副突变(paramutation)现象可以被多代遗传,也是DNA是遗传的分子基础这一通用法则的极少数例外。
遗传变化
突变

在DNA复制的过程中,第二链的聚合中偶尔会产生复制错误。这些错误被称为突变,它们能够对于一个生物体的表现型产生影响,特别是当它们位于一个基因的蛋白质编码区中时。错误率通常非常低:每0.1 - 1亿个碱基才会出现1个错误;这是由于DNA聚合酶具有“校对”能力。 没有校对机制则错误率会增加1000倍,例如许多病毒所依赖的DNA或RNA聚合酶缺乏校对能力,这使得病毒复制过程具有很高的突变率。能够增加DNA发生改变的几率的因素被称为突变原:一些化学品常常可以通过影响正常的碱基对结构来提高DNA复制中的错误率,而紫外线能通过破坏DNA结构来诱发突变。 由于对DNA的伤害在自然界中随时都会发生,细胞则利用DNA修复机制来修复DNA中存在的错误配对和断裂,但有时也无法将受破坏的DNA还原到破坏前的序列。
在利用染色体互换来交换DNA和重组基因的生物体中,减数分裂过程中所出现的配对错误也会导致突变。 当相似序列导致姐妹染色体产生错误配对时,这种染色体互换出现错误的可能性非常大;这使得基因组中一些区域更趋向于以这一方式发生突变。这些错误能够对DNA序列产生很大的结构变化:整个区域的重复、倒位或刪除,或者不同染色体之间发生意外性的交换(被称为染色体易位)。
自然选择与进化

突变会使生物体具有不同的基因型,并可能导致不同的表现型。许多突变对于生物体的表现型、健康和繁殖适应性基本没有影响。有影响的突变则往往是有害的,但也有少量是有益的。在对果蝇的研究中发现,如果一个突变改变了基因所编码的蛋白质,那这一突变很可能是有害的(大约有70%的此类突变具有破坏性影响,而剩余的突变则是中性的或微弱有益的)。
群体遗传学是研究基因变异在生物群体中的分布和这一分布的时间变化性。 一个群体中的一个等位基因的变化频率会受到自然选择的影响,具有更高的存活率和繁殖率的等位基因能够随着时间而越来越频繁地出现在该群体中。 此外,遗传漂变能够引发等位基因出现频率的随机变化而不受自然选择的影响。
在经过多个世代的传承后,生物体的基因组会发生改变,引起被称为进化的现象。突变和对于有益突变的选择使得一个物种不断地进化到能够更好地在所处的环境中生存下来的形式,这一过程被称为适者生存。新的物种的形成常常是由于地理分离而造成的,地理上的分离使得不同种群能够在遗传学上独立发展而产生分化。 遗传学定律在群体生物学和进化研究中的应用被总结为现代综合理论。
由于进化过程中的序列分化和变化,物种的DNA序列之间的差异可以用作“分子时钟”(molecular clock)来计算物种之间的进化距离。 遗传比较被普遍认为是鉴定物种之间亲缘关系的最准确的方法,过去常用的方法则是比较物种之间的表现型的特征。物种之间的进化距离可以用进化树来综合表示,进化树可以表示由共同祖先随时间分化而来的物种之间的亲缘关系,但不能表示无亲缘关系的物种之间的遗传物质的转移(被称为基因水平转移,在细菌中非常普遍)。
研究与技术
模式生物与遗传学

一开始遗传学家们的研究对象很广泛,但逐渐地集中到一些特定物种(模式生物)的遗传学上。这是由于新的研究者更趋向于选择一些已经获得广泛研究的生物体作为研究目标,使得模式生物成为多数遗传学研究的基础。 模式生物的遗传学研究包括基因调控以及发育和癌症相关基因的研究。
模式生物具有传代时间短、易于基因操纵等优点,使得它们成为流行的遗传学研究工具。目前广泛使用的模式生物包括:大肠杆菌(Escherichia coli)、酿酒酵母(Saccharomyces cerevisiae)、拟南芥(Arabidopsis thaliana)、线虫(Caenorhabditis elegans)、果蝇(Drosophila melanogaster)以及小鼠(Mus musculus)。
医学相关的遗传学研究
医学遗传学的目的是了解基因变异与人类健康和疾病的关系。 当寻找一个可能与某种疾病相关的未知基因时,研究者通常会用遗传连锁和遗传系谱来定位基因组上与该疾病相关的区域。在群体水平上,研究者会采用孟德尔随机法来寻找基因组上与该疾病相关的区域,这一方法也特别适用于不能被单个基因所定义的多基因性状。 一旦候选基因被发现,就需要对模式生物中的对应基因(直系同源基因)进行更多的研究。对于遗传疾病的研究,越来越多发展起来的研究基因型的技术也被引入到药物遗传学中,来研究基因型如何影响药物反应。
癌症虽然不是传统意义上的遗传病,但被认为是一种遗传性疾病。 癌症在机体内的产生过程是一个综合性事件。机体内的细胞在分裂过程中有一定几率会发生突变。这些突变虽然不会遗传给下一代,但会影响细胞的行为,在一些情况下会导致细胞更频繁地分裂。有许多生物学机制能够阻止这种情况的发生:信号被传递给这些不正常分裂的细胞并引发其凋亡;但有时更多的突变使得细胞忽略这些信号。这时机体内的自然选择和逐渐积累起来的突变使得这些细胞开始无限制生长,从而成为癌症性肿瘤(恶性肿瘤),并侵染机体的各个器官。
相关研究技术

可以在实验室中对DNA进行操纵。限制性內切酶是一种常用的剪切特异性序列的酶,用于制造预定的DNA片断。 然后利用DNA连接酶将这些片断重新连接,通过将不同来源地DNA片断连接到一起,就可以获得重组DNA。重组DNA技术通常被用于在质粒(一种短的环形DNA片断,含有少量基因)中,这常常与转基因生物的制造有关。将质粒转入细菌中,再在琼脂平板培养基上生长这些细菌(来分离菌落克隆),然后研究者们就可以用克隆菌落来扩增插入的质粒DNA片断(这一过程被称为分子克隆)。
DNA还能够通过一个被称为聚合酶链锁反应(又被称为PCR)的技术来进行扩增。 利用特定的短的DNA序列,PCR技术可以分离和扩增DNA上的靶区域。因为只需要极少量的DNA就可以进行扩增,该技术也常常被用于DNA检测(检测特定DNA序列的存在与否)。
DNA测序与基因组
DNA测序技术是遗传学研究中发展起来的一个最基本的技术,它使得研究者可以确定DNA片段的核苷酸序列。由弗雷德里克·桑格和他的同事于1977年发展出来的链终止测序法现在已经是DNA测序的常规手段。 在这一技术的帮助下,研究者们能够对与人类疾病相关的DNA序列进行研究。
由于测序已经变得相对廉价,而且在计算机技术的辅助下,可以将大量不同片断的序列信息连接起来(这一过程被称为“基因组组装”),因此许多生物(包括人类)的基因组测序已经完成。 这些技术也被用在测定人类基因组序列,使得人类基因组计划得以在2003年完成。 随着新的高通量测序技术的发展,DNA测序的费用被大大降低,许多研究者希望能够将测定一个人的基因组信息的价格降到一千美元以内,从而使大众测序成为可能。
大量测定的基因组序列信息催生了一个新的研究领域——基因组学,研究者利用计算机软件查找和研究生物的全基因组中存在的规律。基因组学也能够被归类为生物信息学(利用计算的方法来分析生物学数据)下的一个领域。
合成生物技术
系统遗传学(System Genetics)的应用 - 合成生物学(Synthetic Biology),后基因组时代,合成生物技术采用计算机辅助设计、全基因与基因组合成等技术,进行细胞的信号传导与基因调控网络的人工设计的基因工程技术,继遗传育种、细胞杂交、转基因生物等生物技术之后,将成为21世纪遗传工程发展的新趋势,系统遗传学揭示基因组的结构、功能与演化的生物系统进化机理,涉及医学遗传学、医学心理学等基因系统调控、信号传导网络等研究,为肿瘤、遗传病、精神病、衰老等疾病发生的诊断与药物筛选、制药产业等开拓了新的途径。
参见
引用
来源
- 书籍
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, and Walter P. Molecular Biology of the Cell 4th edition. 2002. ISBN 0-8153-3218-1 (英语).
- Griffiths AJF, Miller JH, Suzuki DT, Lewontin RC, and Gelbart WM. An Introduction to Genetic Analysis. New York: W.H. Freeman and Company. 2000. ISBN 0-7167-3520-2 (英语).
- Hartl D, Jones E. Genetics: Analysis of Genes and Genomes 6th. Jones & Bartlett. 2005. ISBN 0-7637-1511-5 (英语).
- Lodish H, Berk A, Zipursky LS, Matsudaira P, Baltimore D, and Darnell J. Molecular Cell Biology 4th edition. 2000. ISBN 0-7167-3136-3 (英语).
延伸阅读
- 戴灼华、王亚馥、粟翼玟. 《遗传学》 第二版. 高等教育出版社. 2008年. ISBN 978-7-04-022083-4 (中文(简体)).
- 李振刚. 《分子遗传学》 第三版. 科学出版社. 2008年. ISBN 978-7-03-020134-8 (中文(简体)).
- Robert H. Tamarin. Principles of Genetics. WCB/McGraw-Hill. 1999. ISBN 0697354628 (英语).
- Susan L. Elrod, William D. Stansfield. Schaum's Outline of Theory and Problems of Genetics. McGraw-Hill Professional. 2001. ISBN 0071362061 (英语).
- Leland Hartwell. Genetics: From Genes to Genomes. McGraw-Hill. 2008. ISBN 978-0072848465 (英语).
外部链接
| 维基共享资源中相关的多媒体资源:遗传学 |
| |||||||||||||||||||||||||

| ||||||||||||||||||||||||||||||||
|