
| 真菌界 化石时期: (詳見「化石紀錄」一節)
| |
|---|---|

| |
| 從左上角起順時針: 擔子菌門的毒蠅傘; 子囊菌門的肉杯菌; 發霉的麵包; 一種壺菌; 麴菌的分生孢子 | |
|
科学分类 | |
| 域: | 真核域 Eukaryota |
| 演化支: | 單鞭毛生物 Amorphea |
| 演化支: | OBA 超類群 Obazoa |
| 演化支: | 後鞭毛生物 Opisthokonta |
| 总界: | 菌物总界 Holomycota |
| 演化支: | 動孢菌 Zoosporia |
| 界: |
真菌界 Fungi (L., 1753) R.T.Moore, 1980 |
| 門 | |
|
(Tedersoo等人2018年發表的分類) | |
真菌界(學名:Fungi)生物通稱真菌,是真核生物中的一大類群,包含酵母、黴菌等微生物,以及為人熟知的菇類。真菌與動物同為異營生物,需依賴其他生物所所產的有機物為碳來源,通常會以滲透營養的方式取得養分,即分泌大量的酶將環境中有機物大分子分解成小分子,再把小分子透過擴散作用吸入到細胞中。真菌的一大特點為以幾丁質為主要成分的細胞壁,和無細胞壁的動物及細胞壁主成分為纖維素的植物截然不同。
真菌自成一界,獨立於植物、動物和其他真核生物;真菌也不能進行光合作用,其成長型態與植物一樣不能移動,但可以透過菌絲的延長拓展棲地,也能透過經由有性或無性生殖產生的孢子進行長距離的傳播(某些孢子還具有鞭毛,可在水中移動)。真菌是生態系中的主要分解者,且在生態系的物質循環扮演重要角色。分子系统发生学顯示真菌在演化上是一個單系群,由一個共同祖先演化而來。黏菌與卵菌(水黴菌)在歷史上曾因形態相似而歸屬真菌界,但分子支序顯示它們與真菌的親緣關係甚遠,其和真菌的相似性是趨同演化的結果。研究真菌的學科稱為真菌學,過去學者們曾因為真菌和植物形態類似,而通常將其視為植物學的一個分支。但分子證據顯示,真菌和動物之間的關係要比和植物之間更加親近,真菌與動物同屬後鞭毛生物,兩者在演化上的關係也是分類學的研究熱點之一。
真菌廣佈於全世界,但多數真菌體型很小,或存在土壤與水域中,常被人所忽略,某些物種會產生稱為孢子果(Sporocarp)的大型子實體,也就是蕈類。真菌可與動物、植物或其他真菌產生互利共生或寄生等交互作用,例如與植物形成菌根,以及與藻類或藍綠菌形成地衣。部分真菌中的生物鹼與聚酮等許多物質具有生物活性的物質,稱為真菌毒素,對包括人類在內的動物有毒。一些物種的孢子含有精神藥物的成份,被用在娛樂及宗教儀式上。真菌可以分解人造的物質及建物,並在人類及其他動物造成疾病,真菌感染或食物腐敗引起的作物損失對人類的食物供給和區域經濟產生很大的影響,而真菌病則包括足癬等淺表層的感染,與隱球菌病等全身感染的疾病,嚴重時甚至可能致死。有些蕈類被人作為食物,有些真菌則是食品工業的要角,可以作為麵包的膨鬆劑,或發酵製造醬油、豆腐乳與酒等食品。1940年代起,真菌亦被用來製造抗生素,現在許多工業所需的酵素也是由真菌所合成。有些真菌可以被當作生物農藥,用來抑制雜草、植物疾病及害蟲。
真菌各門的物種之間不論是在生態、生活史及形態(從單細胞水生的壺菌到巨大的菇類)都有很巨大的差別。人類對真菌各門真正的生物多樣性了解得很少,預估約有220萬-380萬個物種,但目前已發表的僅有12萬種。自從18、19世紀,卡爾·林奈、克里斯蒂安·亨德里克·珀森及伊利阿斯·馬格努斯·弗里斯等人在分類學上有了開創性的研究成果之後,真菌便已依其形態(如孢子顏色或微觀構造等特徵)或依生理學給予分類。二十世紀分子遺傳學與生物技術的飛速發展下,DNA定序開始被廣泛用於真菌分類,使傳統分類系統受到一定程度的挑戰。最近十幾年來在系统发生学上的研究已多次改變人們對真菌分類的觀點,也有原本不被認為是真菌的生物(微孢子蟲、隱真菌門、核形蟲)被歸入真菌界或與其關係密切的類群中。各類群真菌的關係仍未有定論,且是現在真菌研究的重點之一。
命名
別稱
在英文裏,真菌界的單數是Fungus,複數是Fungi或Funguses,這三者皆可指代真菌。
在中文裏,在科學界也可稱為“菌物界”,在食品領域也可寫為“真蕈”。
詞源
英文及拉丁文
真菌的英文「fungus」直接源自於拉丁文對蘑菇的稱呼,古羅馬作家老普林尼與賀拉斯的作品中都有這個稱呼,而拉丁文的「fungus」則是來自希臘文的海綿「σφογγος」(sphongos),因蘑菇與黴菌形態上的特徵而得名。此一字根也出現其他歐洲語言中,例如德文意指黴菌的「Schimmel」也源於此字根。
真菌學是以科學方法研究真菌的學門,而真菌學的英文「mycology」則是直接來自希臘文中對蘑菇的稱呼「μύκης」(mykes)。「mycology」一詞最早出現於英國自然學家邁爾士·約瑟·貝克萊的著作《English Flora of Sir James Edward Smith》第五卷 。一起存在於某個地理區域的所有真菌合稱真菌相(mycobiota)。
真菌(Fungi)的命名和其所指稱的分類元相當複雜。在1969年羅伯特·魏泰克提出五界說之前,生物學家已發現卵菌和黏菌等並不屬於真菌界的核心成員。1959年魏泰克在處理真菌界時,已經將這些生物劃歸此界的核心之外。於此時期在討論真菌類的指稱時,開始出現所指涉的名詞是指「廣義真菌」或「狹義真菌」(僅包含真菌的核心成員)的問題。1981年,湯瑪斯·卡弗利爾-史密斯曾建議以「真真菌界」(Eufungi)作為狹義真菌的界名。1992年大衛·萊斯里·哈克斯沃思則建議以「fungi」視為廣義真菌的指稱,同年唐納德·J·S·巴爾(Donald J. S. Barr)等人又建議以「總真菌」(Union of Fungi)代指廣義真菌,「Eumycota」則訂為狹義真菌的界名。因此當時在使用Fungi一詞時,必須同時說明指的是廣義真菌還是狹義真菌。
為了解決這個混亂的命名問題,1993年卡弗利爾-史密斯規範真菌界的名稱為「Fungi」,不屬於真菌界核心成員的物種直接畫出真菌界的範圍,不再使用「Eumycota」或「Eufungi」等名詞,目前「真菌界」在共識上稱為「Kingdom Fungi」。黏菌、卵菌,及絲壺菌等非屬核心真菌的物種,近幾年已經明確移出真菌界的範圍內。
中文
中文名稱的脈絡也同樣複雜,中国大陆所說的「真菌」一般慣指「Fungi」;台灣经常以「菌類」甚至代表蘑菇的“蕈类”代指廣義上的真菌。1991年,中国大陆植物病理學家裘維蕃認為比起「真菌界」,採用與「動物界」和「植物界」相對等的「菌物界」更為理想,並將真菌畫為菌物界底下的一個門,因此「菌物」一詞指涉的為廣義真菌。台灣的吳聲華博士則認為現今的「真菌界」指的為狹義真菌。2009年,Liu等人提出菌物總界的概念,含括真菌界與核形蟲等生物,組成了單系性的分類元。
特徵

在分子系统发生学問世以前,分類學家一般認為真菌屬於植物,這是因為真菌與植物有許多共同特徵,例如兩者皆不會移動,大致的形態與棲地也相當類似,真菌與植物都生長在土壤中,且部分真菌形成的產孢結構子實體外型與苔蘚植物相當類似。現在真菌被認為是獨立的一界生物,與植物與動物均不同,它們早在十億年前(大約是新元古代)就在演化上與動物分道揚鑣了。真菌多數的特徵都可見於其他生物中,因此很難找到某項特徵為真菌獨有而不存在其他生物中。真菌與其他生物共同擁有的特徵如下:
- 與所有其他真核生物:真菌有細胞核,基因組中有大量非編碼DNA,基因中也有內插子存在。真菌也具有許多有膜胞器,包括粒線體、內質網、高基氏體等,且膜中含有固醇(但真菌膜中的固醇種類大多為麥角固醇,與動植物膜中的固醇種類不同)。真菌的核糖體大小與其他生物同為80s,並生產許多醣類,包括糖醇(如甘露醇)、雙醣(如海藻糖)與多醣(如也存在於動物體內的肝糖)。作為真核生物。真菌也可以甲羥戊酸與焦磷酸鹽為原料合成萜烯類化合物。
- 與動物:真菌與動物都沒有葉綠體,兩者同為異營生物,都需要以有機物作為碳源。
- 與植物:真菌與植物都具有細胞壁與大型液胞,都可行無性生殖或有性生殖,且真菌與苔蘚、蕨類等許多演化上較早分支的植物都可以產生孢子。真菌與苔蘚的細胞核一般都是單倍體。某些真菌合成的次級代謝產物與植物合成的相當類似,但合成同種次級代謝產物的植物與真菌酵素的蛋白質序列通常大相徑庭,表示這些酵素是分別在植物與真菌中獨立演化而來。另外,大部分真菌都沒有類似維管束的構造可以長距離輸送水分與無機鹽,少數真菌如蜜環菌屬的菌絲會形成中空的管狀結構菌絲索,外型類似植物的根。
- 與裸藻與某些細菌:高等真菌、眼蟲與某些細菌都以一種特殊的生化反應:α-氨基己二酸途徑來合成離胺酸,其他生物則採用二氨基庚二酸途徑。
- 與卵菌:許多真菌與卵菌的細胞可以形成管狀可延伸的構造,稱為菌絲,菌絲中可能有多個細胞核,菌絲的頂端會聚集許多含有脂質、蛋白質等有機物質的囊泡,這些囊泡合稱頂體。不過有些外型相似的生物,例如綠藻,是靠細胞分裂來生長延長。另外也有許多真菌是單細胞生物,不形成菌絲,通稱為酵母菌。也有些真菌(雙態性真菌)同時具有酵母菌與菌絲兩種形態。
- 類似某些動物與植物,有超過70種真菌可以發光。
多樣性

真菌廣泛分佈於全世界的各種棲地中,包括氣候極端的環境如沙漠、深海以及受輻射污染的地區 ,有些種類對輻射的抗性極強,某些地衣甚至可以抵擋高強度的宇宙射線與紫外線,在人造衛星上的研究站存活。大多數真菌生長在陸地上,但有些真菌生活史的部分或全部階段生長於水中,例如造成全球兩生類數量下降的蛙壺菌,壺菌門的真菌會產生在水中游動的動孢子以尋找兩生類作為宿主。還有一些真菌生活在海洋中。
目前被生物分类学家们正式记录与描述的真菌种类约为12万种,但人們對於世界真菌多樣性的了解仍相當有限。一項2017年的研究估計真菌種類應該介於220萬至380萬之間。歷史上有許多方法可以用來鑑定真菌物種,傳統真菌學主要以孢子或子實體的大小、形狀等形態上的特徵分辨真菌,另一種鑑定物種的方法是透過生理與生化性質,即依真菌的化學測試的結果來分辨。分子生物學興起後,DNA測序與支序分析等技術對真菌多樣性的研究起了決定性的作用,不但大大提高了對新物種的分辨能力(特別是隱真菌門等傳統上不清楚的低等真菌),也更能反映不同類群真菌在演化上的關聯。
真菌學

真菌學是生物學的一個分支,研究真菌的遺傳、生化性質、分類、對人類的用途(包括藥用、食用與宗教致幻劑)以及對人類的威脅(包括毒性與感染)。因為許多植物病原是真菌,植物病理學的研究也常與真菌學高度相關。
人類對真菌的利用可以追溯到史前時代,早在5300年前的新石器時代,在奧地利阿爾卑斯山區發現的木乃伊冰人奧茨遺骸上便帶有兩種多孔菌,其中木蹄層孔菌可能是用來生火的燃料,白樺菌菇則可能是藥用。古代人類常在不知情的狀況下以真菌為食物來源,例如發酵麵包與果汁的生產都有真菌參與。也有早期的手寫記錄描述作物遭到疑似為真菌的病原感染而破壞。
歷史
真菌學是一門相對較新的科學,在十七世紀顯微鏡發明後才出現。早在1588年,吉安巴蒂斯塔·德拉·波尔塔就發現了真菌的孢子,但真菌學真正的濫觴一般認為是1729年義大利植物學家皮耶爾·安東尼奧·米凱利出版的《植物新屬》,他除了觀察到孢子之外,還發現在適當條件下孢子可以萌發,並生長成當初產生該孢子的同一種真菌。荷蘭科學家克里斯蒂安·亨德里克·珀森被認為是現代真菌學的開創者,他以林奈的二名法為基礎建立了第一套菇類分類基礎。隨後伊利阿斯·马格努斯·弗里斯進一步採集了大量真菌標本,並以孢子顏色與其他微觀結構對真菌進行分類,許多標準至今仍被分類學家採用。其他重要的真菌學者還包括邁爾士·約瑟·貝克萊、奧古斯特·卡爾·約瑟夫·科爾達、漢瑞其‧安東‧狄伯瑞、路易·勒內·圖拉斯內與查爾斯·圖拉斯內兄弟、亞瑟·亨利·雷金納德·布勒、柯提斯·蓋茨·洛依德與皮尔·安德烈·萨卡多等。二十世紀隨著分子生物學、生物化學、遺傳學與生物技術的突破,真菌學的研究方法逐漸現代化,DNA定序與支序分類學為真菌多樣性與演化的研究打開了另一扇門,也挑戰了許多傳統以形態為基礎的分類系統觀點。
形態
微觀結構

許多真菌以菌絲的形式生長,菌絲是直徑2-10微米的圓柱體絲狀結構,可長達數公分。菌絲一般自其頂端往前生長,新的菌絲形成通常是舊有菌絲的中間產生了一個新的生長點,而長出新的分支,也有些情況是生長中的菌絲頂端分岔成兩條菌絲。菌絲成長後相互交錯,成為真菌的營養構造菌絲體。菌絲可以是多核(無隔板)或是有隔板的,有隔板的菌絲不同細胞核間有細胞壁將其分隔,使一個隔間具有一或數個細胞核,多核菌絲則沒有隔板,基本上就是一個含有多個細胞核的巨大細胞。隔板上通常有小孔可以使細胞質與胞器流通,有時細胞核也可以通過小孔,例如擔子菌門真菌形成的桶孔隔膜 。
許多真菌的菌絲形成特化的構造,以更容易從宿主身上吸收養分,例如許多植物病原真菌具有吸器,許多球囊菌門的真菌則形成叢枝(Arbuscular)穿透共生的植物細胞以吸取養份。
雖然真菌屬於後鞭毛生物,其共祖有一根後生的鞭毛,但除了壺菌門以及原本屬於壺菌門的新美鞭菌門和芽枝黴門外,其他真菌都已經失去了鞭毛。真菌的另一項特徵是多數種類具有以幾丁質與葡聚糖構成的細胞壁。
巨觀結構

真菌的菌絲常是肉眼可見的,例如在發黴的牆壁或腐敗的食物上,這種菌絲體俗稱黴菌,而長在實驗室培養基上的菌絲則一般稱為菌落,菌落的形狀與顏色常是鑑定真菌的重要指標。有些真菌相當長壽,且可以長到非常大的體積,美國馬盧爾國家森林的一個蜜環菌菌落已生長到超過900公頃,其生長時間推測已超過9000年。
子囊果與擔子果分別是子囊菌門與擔子菌門真菌的子實體,內含可以產生有性孢子的子實層。
生長與生理

真菌在固體介質中以菌絲形式生長,在水中則常以單細胞酵母菌形式生長,皆是為了提升表面積以增加吸收養分的效率。菌絲特別適合在固體介質上生長,且可以形成較大的穿透力而適於侵入新的固體介質,例如稻熱病菌等植物病原菌特化出壓器的構造以穿透植物組織,壓器可藉由增加細胞中甘油等溶質的濃度以加大膨壓,在植物表皮上產生高達1200psi的壓力。淡紫擬青黴菌也用一種類似的結構來穿透線蟲的卵。
絕大多數菌絲都只能自頂端往前生長,Neotyphodium等部分內生真菌則是少見的例外,它們的菌絲附著於宿主細胞上,並可能因宿主細胞延長造成的機械拉力,而促使其菌絲從中間分裂生長(intercalary extension),以長至跟宿主的組織一樣長度。另外擔子菌子實體形成時,菌柄等大型器官的長成是菌絲體積膨大的結果。真菌形成多細胞組織有數種功能,例如子實體是為了產生與散播有性孢子,生物膜則有助於佔據基質與進行細胞溝通,多細胞個體的出現在真菌、動物與植物中都是各自獨立演化出來的。
真菌傳統上被認為是異營生物,只能依賴其他生物固碳形成的有機碳為碳原,演化造成真菌的代謝能力相當多樣化,可以分解各式各樣的有機物,有些種類甚至可能使用黑色素從伽馬射線等輻射中獲取能量,目前只知道少數真菌具有以輻射為能源進行生長的能力,背後的生理機制也不甚清楚,不過整個過程可能與植物利用可見光行光合作用固碳類似,但採用輻射能作為能量來源。
繁殖

真菌的繁殖機制相當複雜,反映了此類生物生活史與遺傳組成豐富的多樣性 。據統計有超過三分之一的真菌物種採用一種以上的方式繁殖,例如無性時期產生無性孢子,有性時期又產生不同種類的有性孢子。不同的環境刺激可以啟動特定基因表現,讓真菌長出不同的特化結構以產生無性或有性孢子 。
無性生殖
真菌的無性生殖有透過產生無性孢子與菌絲斷裂等機制,前者包括以孢子囊、分生孢子囊柄等特化的結構產生孢子,後者則是菌絲斷裂成為許多片段,每一片段都長成一條新的菌絲。無性生殖可以產生基因上與母體相同、佔據同樣生態棲位的子代,繁殖效率也較有性生殖迅速,可以在短時間快速拓殖。不完全菌是一個舊時的分類名詞,指未觀察到有性世代的真菌,現在分類學上已不使用此名詞,但仍可以用來泛指缺乏有性世代的真菌。
有性生殖
除了球囊菌門與微孢子蟲外,其他門的真菌都可透過減數分裂產生配子、進行有性生殖(且基因組分析顯示球囊菌門與微孢子蟲的真菌應該也能進行有性生殖,只是尚未觀察到)。真菌有性生殖的過程與動物、植物相當不同,而不同真菌間特化來進行有性生殖的結構也有差異,是鑑定真菌物種的重要依據,而拿不同真菌進行交配實驗,觀察兩者能不能形成子實體,可以透過生物物種概念(biological species concepts)以兩者能不能成功交配、交配產生的子代有沒有生殖能力來判斷它們是不是同一物種 。主要的生物類群早期都是以有性生殖特化的結構與孢子差異來描述、區分的,例如產生子囊的子囊菌門與產生擔子的擔子菌門。有一些物種還有交配型之分,必須不同交配型的兩個體才可以交配(heterothallic),其他物種則可以和所有個體交配,甚至自交產生孢子(homothallic)。
許多真菌的生活史均存在單倍體與二倍體的時期,真菌進行有性生殖時,菌絲的融合形成交錯的網絡,這個現象稱為融合(anastomosis),是有性生殖不可或缺的。許多子囊菌與擔子菌接著會進入雙核體的時期,即菌絲中一個細胞裡有兩個細胞核,分別來自兩個親代個體,這兩個細胞核不會馬上進行核融合(karyogamy),而會在細胞中共存一段時間。子囊菌門子实层中雙核的菌絲會形成一個鉤狀結構(crozier),以確保細胞核分裂時,兩個子細胞核可以分別妥善分配到上方與下方的細胞中,維持菌絲異質雙核(有來自雙親的兩個細胞核)的狀態。細胞中的兩個核融合之後,會立刻進行減數分裂產生子囊孢子,子囊孢子釋放、散播到其他地方後可以萌發長成新的單倍體菌絲。擔子菌門真菌的有性生殖過程與子囊菌門的類似,也是始於兩株真菌單倍體的菌絲融合而產生雙核體的菌絲,不過雙核體在擔子菌門更為普遍,除了子實體外,有時連營養菌絲也是雙核的狀態。擔子菌門的子實層會形成一種稱為扣子體的微觀結構,與子囊菌門的鉤狀結構類似,可以控制細胞核分裂後兩個子細胞核的分佈,維持菌絲異質雙核的狀態。擔子菌的子實體會特化出擔子,擔子中的兩個細胞核融合之後進行減數分裂,會產生擔孢子。最為人所知的擔子菌便是菇類,不過擔子菌也能以其他形態存在。
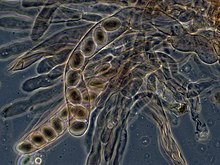
接合菌門的真菌在兩個體的單倍體菌絲融合後,會形成一個特化的孢子囊,孢子囊會進一步特化行成厚壁的接合孢子,對環境的抗性很大,當環境有利真菌生長時接合孢子便會萌發,進行減數分裂以產生單倍體的子細胞,並生長成菌絲。
孢子的釋放

真菌有許多主動將無性與有性孢子釋放的機制,以確保孢子可以離開子實體或菌絲而長距離傳播。許多菌絲生理、機械的機制,以及孢子表面的結構都有助於孢子的散播。某些子囊菌會增加子囊中部分溶質、離子的濃度以提升膨壓,造成水分流入而將子囊漲破,使子囊孢子得以釋出。擔子菌門的擔孢子釋放則與孢子表面水珠造成的表面張力有關,造成孢子彈射出的初始加速度可以高達10,000g 。孢子最終彈射的距離約為0.01-0.02公分,已足以離開蕈褶,進入空氣中。有些真菌孢子的釋放機制較為特殊,馬勃與鳥巢菌的孢子需要雨水等外力將子實體爆破後才能釋出,鬼筆科的真菌甚至具有鮮豔的顏色與特殊的氣味,以吸引昆蟲協助散播孢子。
其他生殖方式
青黴菌與麴菌有準性生殖的現象,兩個個體的菌絲可能發生融合,造成菌絲中存在來自不同個體的細胞核,這些細胞核可能進一步發生核融合,同源染色體間產生基因重組後再分裂形成單倍體的細胞,其過程與有性生殖類似,但沒有減數分裂的發生,其發生頻率與重要性也仍不清楚。除了在同種真菌發生外,準性生殖還可能在不同種真菌間發生,並可能是真菌演化的機制之一。
化石紀錄
許多真菌的結構都是微觀的,而主要巨觀的結構如子實體又非常柔軟且容易分解,因此與動植物相比,真菌的化石記錄相當稀少,且真菌化石很難與其他微生物的化石分別,除非有現生真菌與化石物種外型相似才較容易分辨。真菌的化石常與動植物的一起出現,通常經過薄切面後,使用光学显微镜或透射电子显微镜檢視觀察,研究壓紋化石時則會使用酸性溶液將化石周圍的基質溶解,再使用光學顯微鏡或扫描电子显微镜觀察表面的詳細構造。
最早具有典型真菌特徵的化石可追溯至24億年前的古元古代,是棲息於海底的生物,具有可能可以互相接合的菌絲狀構造 。另有研究比較相關生物類群的演化速率,以分子時鐘推測真菌應該在7-10億年前出現。古生代時許多真菌都是水生的,且像現生的壶菌一樣具有鞭毛,但也有部分真菌在寒武纪時就開始往陸地生長,較陸生植物的出現早上許多。隨後真菌演化出許多陸地生活所需的特徵,包括寄生、腐生、菌根與地衣等與其他生物的交互作用。2009年的一篇研究顯示子囊菌門的共祖便是腐生的,隨後多次在不同類群中,獨立演化出地衣的生長型態。有奥陶纪的真菌化石出現菌絲、孢子等特徵,和現生的球囊菌門真菌相當類似,當時陸生植物仍只有許多類似現生苔蘚的種類,尚不具有維管束 。原杉菌是一種重要的真菌化石,出現於志留紀晚期,並可能是當時陸地上高度最高的生物。泥盆紀早期真菌化石的種類開始變多,且特徵較更早化石典型許多,對其是否屬於真菌爭議漸漸減少,許多壺菌門與接合菌門的化石在萊尼燧石層中被發現。擔子菌門與子囊菌門也差不多在此時出現,到石炭紀的賓夕法尼亞世,多數現生真菌的綱都已經出現了。
形態類似地衣的化石最早在5-6億年前的陡山沱地層中就出現了,地衣是早期陸域生態系的重要成員,最早確定為地衣的化石出現於4億年前,約與最早的子實體化石同期。具有與現生擔子菌扣子體相似結構的化石也出現於石炭紀賓夕法尼亞世的地層,與一株蕨類的化石一起出現 。同擔子菌亞綱的真菌化石則相對較少,兩枚保存於琥珀中的標本顯示白堊紀晚期(9000萬年前)已有外型類似菇類的真菌出現。
二叠纪-三叠纪灭绝事件之後不久,真菌化石大量出現(P-T fungal spike),孢子也大量出現在岩層中,顯示真菌當時是相當佔優勢的生物,這個時期的化石幾乎全部都是真菌化石,但也有研究反對這項理論,認為此一爆發並沒有發生在世界所有地區,某些地區發現的真菌化石爆發和滅絕事件時間不完全吻合,且不易區分這些孢子是來自真菌或藻類。
6550萬年前,許多動植物在白垩纪-第三纪灭绝事件中滅絕,緊接著真菌再度大量出現,可能是世界各地森林大面積毀滅後,為真菌提供腐生生長的環境造成。而真菌的拓殖甚至可能助長了哺乳類在新生代的繁盛,因為哺乳類相對於其他脊椎動物,對真菌感染的抗性特別大。
分類
真菌學在傳統上常被劃歸在植物學的範疇中,但真菌其實與動物的親緣更為接近。分類學家根據分子系统发生学的分析,將廣義的真菌與動物(菌物總界與動物總界)共同組為稱為後鞭毛生物的單系群,而真菌界本身也是一個單系群。真菌的分類經常有所變動,特別是DNA定序技術普及後,越來越多研究比較不同真菌間的DNA序列,不斷提出新的分類觀點。許多傳統上以形態或交配實驗所做出的分類系統受到相當大的挑戰。
目前真菌較高層級的分類系統仍有很大爭議,新理論不斷被提出,各個分類階層的名稱均常有變動。且同一種真菌還可能在生活史的不同階段,例如無性與有性世代擁有數種不同的學名,使真菌分類更加複雜。目前有Index Fungorum與ITIS等資料庫記錄了所有現生真菌的名稱(包含舊分類系統下的同物異名在內),許多分類學家也正努力建立更一致的命名系統 。
2007年,在數十位真菌學家與分類學家通力研究之下,誕生了一套新的真菌分類系統,其中子囊菌門與擔子菌門共同組成雙核亞界,是真菌中多樣性最高的類群,包含絕大多數蕈類、植物病原菌以及用於食品工業的真菌,而傳統分類系統中的壺菌門與接合菌門都被認為是並系群而有所調整,芽枝黴門與新美鞭菌門從壺菌門中分出,成為兩個獨立的門,接合菌門則被分拆成球囊菌門與毛黴亞門、蟲黴亞門、梳黴亞門和捕蟲黴亞門等四個亞門。2018年,Tedersoo等人又發表了新的真菌分類系統,將真菌細分至十八個門。下面的演化樹是根據Silar等人(2016年)與Tedersoo等人(2018年)做出的新分類系統繪製,包含真菌的主要分類單元。注意演化支的長度與演化距離無關: (页面存档备份,存于互联网档案馆)
| 真菌界 |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 担子菌门 |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 子囊菌门 |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
分類類群

隨著分子系统发生学的問世,以DNA序列為基礎的分類方式漸漸取代傳統分類系統,微孢子蟲等許多在五界說被歸屬於原生生物的微生物被發現與真菌的親緣關係較近,而被歸入真菌界中。隱真菌門的物種是土壤與水中的微生物,它們缺乏細胞壁,可以像動物般以吞噬作用取得養分,DNA序列分析顯示它們與微孢子蟲關係非常密切,後者可能是隱真菌門中一個特別深的演化支,兩者共同構成其他真菌的姊妹群,也被劃入真菌界中。另外核形蟲與泉生蟲也與真菌的親緣關係接近,兩者構成包含隱真菌門在內所有真菌的姊妹群,也有研究者將它們劃入真菌界 ,但因兩者缺乏許多真菌共有的特徵,生活型態也與真菌差異較大,Terdersoo等人2018年發表的分類系統選擇將它們獨立成一個界:核形蟲界(Nucleariae)。核形蟲界雖不屬於真菌界,但因與真菌親緣關係密切,是廣義的真菌:真菌總界的成員。
2007年的分類系統將真菌分為微孢子蟲門、壺菌門、芽枝黴門、新美鞭菌門、子囊菌門、擔子菌門以及原屬接合菌門的球囊菌門與毛黴亞門、蟲黴亞門、梳黴亞門和捕蟲黴亞門等四個亞門。2018年Tedersoo等人發表的分類系統則總共將真菌分為九大類群,共計十八個門。包括隱真菌門、Aphelidiomycota、芽枝黴門、壺菌門、單毛壺菌門、新美鞭菌門、油壺菌門、蛙糞黴門、捕蟲黴門、梳黴門、蟲黴門、Calcarisporiellomycota、毛黴門、被孢黴門、球囊菌門、根腫黑粉菌門、子囊菌門與擔子菌門。此次分類所作出較大的改動包括認定微孢子蟲屬於隱真菌門、將壺菌門進一步分拆、將蛙糞黴從蟲黴菌中獨立成一門、將毛黴門進一步分拆等。
微孢子蟲是一種單細胞生物,在動物細胞內行絕對寄生,因為基因組與胞器都高度退化,一度被認為是原始的真核生物。不過DNA序列分析顯示微孢子蟲屬於真菌,而且是所有其他真菌的姊妹群,是真菌演化上最早分支的演化支。近期還有研究認為微孢子蟲與隱真菌門關係密切。
壺菌與現在已獨立出去的新美鞭菌門、單毛壺菌門、芽枝黴門,以及新加入真菌界的隱真菌門與Aphelidiomycota等類群具有動孢子,動孢子具有鞭毛而可以在水中移動,因此壺菌早期曾被歸為原生動物,後來rDNA序列分析顯示壺菌屬於真菌,而且也是真菌演化上較早分支的類群。
芽枝黴門以前認為是壺菌的一支,不過新研究顯示芽枝黴門不屬於壺菌,且可能是雙核亞界與原屬接合菌門的所有真菌的姊妹群,在演化上比壺菌晚分支出去。芽枝黴門多為腐生,以分解有機質為生,其成員可寄生於許多真核生物。與壺菌不同的是芽枝黴門的生活史存在世代交替。
新美鞭菌門也曾被認為是壺菌的一支,其成員是厭氧生物,棲息於大型草食動物的消化系統中 ,它們缺乏粒線體,但有粒線體衍生而成的氢酶体。新美鞭菌門的動孢子也具有鞭毛,還有些種類具有多根鞭毛。


過去屬於接合菌門的生物現在被分拆成數個不同的門,這些真菌包括會造成麵包發黴的黑根黴、可用來製作豆腐乳的毛黴與根毛黴。球囊菌門的真菌會侵入植物的根部,與植物形成叢枝菌根,可幫助植物吸收土壤中的水分與無機鹽,叢枝菌根在演化上出現的時間相當早,最早可追溯至四億年前,這種互利共生可能對陸生植物早期在陸地的適應非常重要。球囊菌門過去屬於接合菌門,2001年首次被提升到門的地位。
子囊菌門是真菌中種類最多的類群,它們可經由減數分裂,在子囊中產生子囊孢子。子囊菌門的真菌包含羊肚菌、松露、多數酵母菌(念珠菌、釀酒酵母、克鲁维酵母属與毕赤酵母)以及某些蕈類,生活型態包括腐生、寄生或與藻類或藍綠菌形成互利共生的地衣。子囊菌門的重要成員還有麴黴屬、青黴菌、麥角菌與鐮孢菌屬等。有一些子囊菌只有觀察到無性生殖,沒有觀察到有性世代的紀錄,但分子序列分析足以顯示其分類屬於子囊菌門。也因為其減數分裂的產物子囊孢子會留在子囊中,而不會馬上散播出去,子囊菌門的真菌(粉色麵包黴菌)曾是遺傳學研究的重要工具。
擔子菌門真菌的菌絲特化出稱為擔子的桿狀構造,以進行減數分裂產生擔孢子。多數蕈類都屬於擔子菌門,另外擔子菌門還包括銹菌與黑粉菌等重要的植物病原菌、存在於人類皮膚上的馬拉色菌屬以及可造成機會性感染的新型隱球菌。
类似真菌的生物
因為形態與生活史的相似,黏菌、卵菌與絲壺菌都曾被認為是真菌界的生物,卵菌、絲壺菌與部分屬於「Phytomyxea」的黏菌,曾與壺菌共同組成真菌界下的鞭毛菌亞門,更早以前還有藻菌、裸菌等分類單元。這些生物現在都已經證實不屬於真菌,真菌的細胞壁是由幾丁質組成,不含有纖維素,卵菌的細胞壁則是纖維素組成的,絲壺菌的細胞壁兼有幾丁質與細胞壁,多數黏菌生活史中的多數階段則是沒有細胞壁的,而是以吞噬作用取得養分,不像真菌、卵菌與絲壺菌是以滲透營養的方式,以擴散作用從胞外吸收養分,唯一的例外是網狀黏菌,這種黏菌具有細胞壁,且也是使用滲透營養的方式獲得養份。現在的分類系統中,卵菌、絲壺菌與網狀黏菌同屬SAR超类群,黏菌則包含屬於變形蟲界、有孔蟲界、古蟲界的物種,與真菌的親緣關係都相當遠,它們相似的外型與生活史只是趨同演化的結果。另外有一種黏菌稱為泉生蟲,與真菌、動物同屬後鞭毛生物,且與核形蟲共同組成現生真菌的姊妹群,與現生真菌關係非常密切。不過關於這些生物的研究有時仍被視為真菌學的範疇,出現在真菌學的課本與論文中。
外毛菌目、變形毛菌目、魚孢菌與珊瑚壺菌原本歸屬接合菌門,後來發現它們屬於動物總界,與現生動物的關係更為密切,與真菌相似的外型也是趨同演化的結果。芽囊原蟲属與曾被認為是一種酵母菌 ,Ellobiopsis曾被認為是一種壺菌,兩者現在都歸屬於SAR超类群。
歷史上放線菌也曾因可以形成菌絲狀的構造而被歸為真菌,但很早就被發現屬於細菌,不是真核生物。
生態
真菌存在於地球上幾乎所有環境中,真菌與細菌同為許多生態系中的分解者,在生物地球化學循環與食物鏈中扮演重要角色。真菌將有機物分解成二氧化碳釋放,使碳元素重新回到食物鏈中,且作為腐生生物,真菌可將氮元素從有機物中分解釋放,對氮循環不可或缺。
共生
真菌與許多生物都有交互作用,其關係包括互利共生、片利共生與寄生。
與植物
真菌與植物形成菌根的共生關係,對植物的生長與許多生態系的維持都相當重要,超過九成的植物與真菌形成菌根,其中許多依賴菌根生存。菌根的演化歷史十分古老,早在4億年前就已出現,可以促進植物對硝酸鹽與磷酸鹽等關鍵無機鹽的吸收,在無機鹽濃度較低的土壤中尤為重要,而植物則將部分由光合作用合成的有機物提供給共生的真菌。菌根還有可能媒介植物與植物之間的物質交換,這種物質交換網稱為菌根網,其中菌異營是一個相當特殊的例子,為某些植物寄生真菌,藉由菌根從真菌體內獲取自己所需的有機物,而本身失去了行光合作用的能力,菌異營中有機物的流向與正常菌根是相反的。有些真菌生長於植物的根、芽、葉等組織中,這些真菌稱為內生真菌,內生真菌與菌根一樣常是植物與真菌間的互利共生,真菌生長於植物體內獲得養份與庇護,並可提高植物對草食動物的抗性以讓植物受益。
與藻類

地衣是真菌與行光合作用的藻類的共生產物,在地衣中,行光合作用的部分稱為共生光合生物(photobiont),而真菌則許多來自子囊菌門、少數來自擔子菌門。地衣存在於五大洲的所有生態系,對成土作用與演替的起始非常重要,且能生長在極端環境中,如極地、山地與半乾旱地區。地衣可以生長在其他生物難以生存的基質,如樹皮、石頭、裸土、落葉、貝殼、藤壺等。與菌根類似,藻類等共生光合生物提供真菌有機物,真菌則提供它們水分與無機鹽。地衣中真菌與共生光合生物的共生關係非常緊密,幾乎可視為一個個體,於原本的兩種生物均有相當差別。約有兩成已發表的真菌(約兩萬種)會形成地衣 。地衣的共同特色包括生長速度緩慢、體積小、壽命長、無機鹽多半來自空氣中、且耐乾旱的程度較生長在同環境的其他光合作用生物高。
與昆蟲
許多昆蟲與真菌存在某些交互作用。有些種類的螞蟻會培養真菌作為它們的食物來源,鞘翅目的粉蠹蟲也會在樹皮上培養真菌食用。某些種類葉蜂的雌蜂會將帶有一種真菌(Amylostereum areolatum)孢子的蟲卵產到樹中,孢子萌發成菌絲後可分解樹中的營養,對葉蜂幼蟲的生長有益。至少有一種無針蜂與紅麴黴屬真菌關係密切,其幼蟲以紅麴菌屬真菌為食,成蟲築新巢時也會將菌種帶入新巢中。有些非洲熱帶莽原的白蟻也會培養真菌來食用。念珠菌與Lachancea屬的酵母菌存在於脈翅目、鞘翅目與蟑螂的消化道中,目前尚不確定它們是否對昆蟲宿主有益。在腐木上生長的真菌對木食性昆蟲相當重要,長期下來真菌可以改變腐木上元素的組成,釋放磷、氮、鐵、鋅、銅、鉀等甲蟲生長的必須元素到腐木中,對木食性昆蟲的生長發育相當重要。另外許多雙翅目眼蕈蚊總科的昆蟲(包括蕈蚋科、角菌蚊科的物種)幼蟲以真菌的子實體或菌根為食。
有些真菌會寄生在特定昆蟲身上吸食牠們直至宿主死亡,如冬蟲夏草。
寄生
作為寄生物、病原菌

許多真菌寄生於植物、動物(包括人類)、甚至其他種真菌中。有些真菌是感染農作物的植物病原菌,例如感染稻米的稻熱病菌、造成荷蘭榆樹病的榆枯萎病菌、造成栗疫病的栗疫病菌、感染各種穀類的鐮孢菌、黑粉菌、鏈格菌與旋孢腔菌属等,其中稻熱病菌還可以從活體營養(biotrophic,寄生在活體植物上)轉為死体营养(necrotrophic,吸收其殺死的植物體為生),這些植物病原真菌造成林業與農業的重大損失。淡紫紫黴菌等食肉真菌的菌絲可以特化產生收縮環(constrictive ring)與黏網(adhesive net)以捕食線蟲。隱真菌門與微孢子蟲等真菌基群的物種大多寄生於其他生物的細胞中,隱真菌門的物種可寄生卵菌、藻類、變形蟲或其他真菌,其模式屬羅茲菌屬便是寄生於芽枝黴門的異水黴屬細胞內。微孢子蟲則是行絕對寄生的微生物,其本身細胞結構與基因體都經過相當程度的退化,連粒線體亦失去功能,完全仰賴昆蟲、甲殼類、脊椎動物等其他動物宿主為生,從宿主細胞中獲取ATP。微孢子蟲也有可能感染人類,造成微孢子蟲病。
有些真菌可以感染人類,造成嚴重的疾病,有些感染若沒有受到妥善治療甚至可能致命。真菌感染造成的疾病包括麴菌病、念珠菌症、球黴菌症、隱球菌病、組織胞漿菌病、足菌腫與副球黴菌病等,免疫缺陷的病人特別容易受到麴黴、念珠菌、隱球菌、組織胞漿菌與肺孢子菌等真菌的感染。另外還有些真菌會造成眼睛、皮膚、指甲與頭髮的淺層感染,造成皮癬菌症與足癬等疾病。真菌的孢子也經常引發人的過敏。
作為被寄生的宿主
真菌寄生蟲(mycoparasitic organisms)泛指可以寄生於真菌的生物,腐黴屬生物(屬於卵菌,並非真菌)便可寄生在真菌體內,而有作為針對真菌的生物防治劑的潛力。真菌本身也可以寄生其他種真菌,例如金孢菌寄生是一種子囊菌門的真菌,但寄生在牛肝菌上。 真菌可以被真菌病毒感染。真菌病毒大多屬於雙股RNA病毒,但也有約三成是正义单链RNA病毒。最著名的真菌病毒是可以感染栗疫病菌的CHV1病毒,最早在毒性較弱的栗疫病菌中被發現,已成功用作生物防治劑,在歐洲成功抑制栗疫病菌的生長,但在美國、中國、日本等地使用時可能因為當地栗疫病菌的多樣性太大,有些菌株不能被病毒感染而效果不彰。2016年已有用基因工程改造的CHV1病毒被製造出來,能感染多數栗疫病菌的菌株,可望在栗疫病病情的控制得到更大成效。
真菌毒素
![(6aR,9R)-N-((2R,5S,10aS,10bS)-5-benzyl-10b-hydroxy-2-methyl-3,6-dioxooctahydro-2H-oxazolo[3,2-a] pyrrolo[2,1-c]pyrazin-2-yl)-7-methyl-4,6,6a,7,8,9-hexahydroindolo[4,3-fg] quinoline-9-carboxamide](http://upload.wikimedia.org/wikipedia/commons/thumb/9/94/Ergotamine3.png/220px-Ergotamine3.png)
許多真菌可以產生對動物或植物有毒的物質,稱為真菌毒素,其中對人類影響較大的包括黴菌在腐敗的食物中產生的毒素,以及有毒蕈類的毒素,例如某些鵝膏菌屬產生的毒傘肽。麥角胺是另一種在人類歷史上相當重要的真菌毒素,食用被麥角菌污染的穀物會導致麥角中毒。黃麴毒素由某些能夠感染穀物、堅果的麴黴屬真菌合成,有很大的肝毒性與致癌性。其他包括赭麴黴毒素、展青黴素、伏馬毒素都可能污染人或牲畜的食物而造成中毒。
真菌毒素是真菌的次級代謝物,許多真菌專門用於合成毒素的生化代謝路徑已被發現,這些毒素可以抑制其他真菌生長或避免被食用而增加真菌自身的適應度。許多真菌的次級代謝物被用來當作藥物,詳見#人類利用一節的說明。
致病機制
有些真菌能感染動植物而造成疾病,不過許多動植物已演化出抵抗真菌感染的機制,對此真菌也出現加以反制的方法,例如增加基因重組與DNA修復以修補被宿主免疫機制破壞的DNA。
玉米黑穗菌是一種植物病原菌,在玉米造成黑穗菌病,不過植物已演化出對抗真菌感染的機制。當感染發生時,植物會迅速產生呼吸爆,即在真菌感染的部位產生大量自由基,而黑穗菌可以啟動由YAP1基因介導的抗氧化的機制加以抵抗,此抵抗機制可保護黑穗菌不受植物的防禦系統破壞,對黑穗菌的有效感染相當重要。另外玉米黑穗菌擁有相當完善的DNA修復系統,其DNA在減數分裂期間發生多次重組,可能可以修補其被宿主自由基破壞的DNA,幫助黑穗菌感染時對抗宿主產生的氧化壓力。
新型隱球菌是一種可以生存在動物或植物體內的擔子菌門酵母菌,可以機會性感染免疫缺陷的病人,它被免疫系統中的肺泡巨噬細胞吞噬後可以在肺中造成感染,有些新型隱球菌可以在巨噬細胞內存活,導致感染的潛伏期,對抗真菌劑的抗性,甚至因此有能力侵犯其他器官。新型隱球菌在巨噬細胞內會啟動抗氧化壓力的基因表現,以避免被巨噬細胞胞內的自由基破壞,另一種抗性機制與減數分裂有關,多數新型隱球菌都是a交配型,a交配型的菌體的細胞核一般是單套,但也可能發生「單核體結實」(monokaryotic fruiting),細胞核分裂後再核融合後形成厚垣孢子,雙套的厚垣孢子可以行減數分裂產生擔孢子 ,此一過程受到DMC1基因(與其他真核生物的RAD51以及細菌的RecA同源)調控,減數分裂中發生的基因重組可能有助其修補被自由基破壞的DNA,而增加與其抗性。
人類利用

人類真菌學是研究人類利用真菌的歷史,以及真菌對人類社會影響的學門。長久以來,人類利用真菌來製造、保存食物,或用作其他用途。在許多國家蕈類養殖與蕈類採集都是重要產業。因為許多真菌可以生產有生物活性的天然產物與抗生素,許多真菌已被開發、或正被開發以大量生產抗生素、維他命、抗癌藥物或降低膽固醇的藥物。近期隨著在真菌進行基因工程的技術發展,使真菌的代謝工程應用成為可能。例如基因修飾後的酵母菌可以用來大量生產許多藥物,且因為酵母菌在培養箱中生長迅速,使生產藥物的效率比從原本的來源提煉還要高。
藥用
當代藥用
許多真菌產生的代謝物可以作為藥物。最著名的例子便是包括青黴素在內的抗生素。在自然界中,真菌或細菌產生的抗生素通常有兩種功用,一是高濃度時作為抑制其他微生物生長的抑制劑,以避免其他微生物與自己競爭資源,二是低濃度時作為物種內或物種間群體感應的訊息傳遞分子 。自然界中產生的青黴素苄青黴素(由產黃青黴菌產生)可以有效殺死的細菌相對有限,但其他各式種類的青黴素可用化學方法合成,以達到更廣泛的殺菌能力,現代使用的青黴素便是半合成的產物,先由真菌合成,再針對需要的特殊性質對其進行化學修飾而得到最終產物。除了青黴素外,由真菌生產的抗生素還包括環孢素(在器官移植時用作免疫抑制劑)、夫西地酸(可用來處理耐甲氧西林金黄色葡萄球菌的感染 )與灰黃黴素(可治療真菌感染)。
另外,可以抑制羥甲基戊二酸單醯輔酶A還原酶,繼而抑制膽固醇合成的他汀類藥物,許多也是由真菌合成的,例如由橘青黴合成的美伐他汀,以及由土麴黴與平菇合成的洛伐他汀。真菌也能生產抑制病毒、癌細胞生長的藥物,例如產於雲芝的抗癌藥物雲芝多醣K已在歐洲與日本等地作為抗癌藥物的佐劑使用,產於香菇的香菇多糖也在日本等國被用在癌症治療中。
傳統醫學


許多真菌是中醫等傳統醫學使用的藥材,中醫用來入藥的真菌包括姬松茸、赤芝與冬蟲夏草等。冬蟲夏草是子囊菌門的真菌中華蟲草寄生於蝙蝠蛾科幼蟲後,死亡的幼蟲與菌絲所組成。最早將冬蟲夏草入藥的地方是西藏與尼泊爾,十五世紀的藏醫醫生舒卡·年姆多吉(Zurkhar Nyamnyi Dorje)在其著作《藥方匯集》中提到冬蟲夏草,是冬蟲夏草用於傳統醫學的最早文字紀錄。中國對冬蟲夏草最早的記載是清朝汪昂在《本草备要》一書中所寫「冬虫夏草,甘平,保肺益肾,止血化痰,止劳咳。四川嘉定府所产者佳。冬在土中,形如老蚕,有毛能动,至夏则毛出土上,连身俱化为草。若不取,至冬复化为虫。」中醫認為冬蟲夏草是名貴滋補的草藥,有「甘平保肺,益腎止血,化痰,已勞嗽」之效。
屬於擔子菌門的桑黃是另一種傳統醫學使用的草藥,中醫、韓醫與日本漢方醫學皆有使用紀錄。早在秦漢時的《神農本草經》就有桑黃入藥的紀錄,中醫認為桑黃有止血與治療婦女病的功效,朝鮮時代的醫書《鄉藥集成方》與《東醫寶鑑》均記載桑黃有如靈丹妙藥,江戶時代的日本也將桑黃當作漢方藥。
食用

食用蕈有些是野外採集的,有些則是商業種植的。西方最常食用的蕈類是洋菇,用在沙拉、湯品與許多其他料理中。許多在亞洲常被種植、食用的蕈類,例如草菇、平菇與金針菇也漸漸傳到西方。許多其他種蕈類可由野外採集來食用或販賣,包括松乳菇、羊肚菌、雞油菇、松露、喇叭菌属與美味牛肝菌等,有些種類在市場上的價格十分高昂,通常是特殊料理中的食材。

生產某些種起司時需要把特定真菌加到凝乳中,以產生特殊的風味或口感,例如斯蒂爾頓芝士與羅克福乾酪上的藍色紋路便是羅克福青黴菌造成的。用於製造起司的真菌都是無毒的,但起司成熟或貯藏時,可能有其他種類的真菌生長於其上,而產生多種真菌毒素,是生產起司時需考慮的因素。
發酵食品
釀酒酵母常用來製作麵包與其他小麥加工食品,許多酵母菌可用來釀酒,米麴菌則可用來發酵大豆,製作醬油、日本酒與味噌,根黴則是製作印尼食品丹貝的重要原料。許多與這些真菌親緣相當接近的麴菌屬真菌會產生對人體有害的毒素,但這些真菌大多是馴化物種,經由育種、人擇選出發酵能力佳,又不會產生真菌毒素的真菌種類用於食品工業中 。另外闊恩牌素肉也是真菌衍生的食品,由金黃色鐮孢發酵而成。
有毒真菌

許多真菌對人類是有毒的,毒性可輕至造成腸胃不適或過敏,重至產生幻覺、器官衰竭甚至致死。有毒的蕈類包括錐蓋傘屬、盔孢傘屬、環柄菇屬與最惡名昭彰的鵝膏菌屬,鵝膏菌屬包含號稱毀滅天使的鱗柄白鵝膏與毒鵝膏,後者常被誤認成可食用的草菇而誤食致死,佔人類因毒菇中毒死亡的比例超過半數 。鹿花菌煮熟後一般可以食用,且被許多人視為美食,但生食則有劇毒,油口蘑甚至在中世紀長期被認為可以食用,且受到某些騎士的喜愛,後來才發現這種蕈類有劇毒,會導致橫紋肌溶解症。毒蠅傘也常造成不致死的中毒,多半是當致幻劑吸食所造成,歷史上毒蠅傘曾被歐洲與亞洲的許多人群拿來吸食,直到現在仍被科里亞克族等西伯利亞東北部的民族在薩滿教儀式上用做致幻性藥品。
未受專業訓練的人很難正確分辨蕈類是否可食,因此專家通常建議將所有在野外發現的蕈類視為有毒的,不要食用。若誤食毒菇出現症狀,應保留剩下的菇與嘔吐物等檢體並立即就醫。
蟲害管理

在農業上,根據競爭排除原則,真菌若可以與有害的細菌或其他真菌競爭資源與空間,甚至寄生這些微生物,抑制有害菌,都可用作生物防治以減少病蟲害。許多真菌可抑制蟎、雜草、線蟲或其他有害於作物的真菌生長,在生物防治上產生了許多應用。蟲生真菌如白殭菌、黑殭菌、被毛孢、擬青黴菌與蠟蚧輪枝菌等皆可抑制害蟲生長而用作生物農藥。Neotyphodium、香柱菌屬等內生真菌生長在植物體內,可以合成生物鹼以避免植物受到草食動物啃食,這種特性也被人利用在放牧場等草地的栽培中,經由植物配種選擇分泌物只對昆蟲有毒性,而對家畜無害真菌。
真菌修復
真菌修復是生物修復的一種,意指以真菌來降解環境中的汙染物。已知某些真菌可以將殺蟲劑、除草劑、消毒劑、礦物雜酚油、甚至長鏈石油分解成二氧化碳與水,還可以吸收環境中的重金屬並儲存於子實體中。有些真菌甚至能用來清理受核污染的土地,核廢料中的貧鈾在環境中容易被氧化,而鈾氧化物可以被植物與細菌所吸收,在食物鏈中層層累積。真菌可以經由生物礦化作用將鈾氧化物固定成磷酸鹽礦物,這種型式的鈾不易被生物所吸收,不會進入食物鏈中對生物造成危害。
模式生物
許多生物學上的重大發現都得益於將真菌用作模式生物的研究,例如一基因一酵素假說便是用粉色麵包黴菌的實驗證實的。釀酒酵母、裂殖酵母與小巢狀麴菌也都是重要的模式生物,長久以來用於有關細胞週期、染色體結構與基因表現調控等真核生物遺傳學與細胞生物學的基礎研究。近年來有更多真菌被當成醫學、植物病理與工業研究的模式生物,例如人類的病原菌白色念珠菌、植物病原菌稻熱病菌與工業上常用來大量生產真核生物蛋白質的嗜甲醇酵母菌。
其他
真菌被廣泛用來生產檸檬酸、葡萄糖酸、乳酸與蘋果酸等化學物質 ,以及工業用的酵素,例如洗衣粉裡的脂酶、用來生產纖維素乙醇和牛仔褲酵素石洗的纖維素酶、以及澱粉酶、蔗糖酶、蛋白酶與木聚糖酶。
參見
註腳
參考書目
- Ainsworth GC. Introduction to the History of Mycology. Cambridge, UK: Cambridge University Press. 1976. ISBN 0-521-11295-8.
- Alexopoulos CJ, Mims CW, Blackwell M. Introductory Mycology. John Wiley and Sons. 1996. ISBN 0-471-52229-5.
- Chandler PJ. A Dipterist's Handbook 2nd. The Amateur Entomologists' Society. 2010: 1–525.
- Deacon J. Fungal Biology. Cambridge, Massachusetts: Blackwell Publishers. 2005. ISBN 1-4051-3066-0.
- Hall IR. Edible and Poisonous Mushrooms of the World. Portland, Oregon: Timber Press. 2003. ISBN 0-88192-586-1.
- Hanson JR. The Chemistry of Fungi. Royal Society Of Chemistry. 2008. ISBN 0-85404-136-2.
- Jennings DH, Lysek G. Fungal Biology: Understanding the Fungal Lifestyle. Guildford, UK: Bios Scientific Publishers Ltd. 1996. ISBN 978-1-85996-150-6.
- Kirk PM, Cannon PF, Minter DW, Stalpers JA. Dictionary of the Fungi 10th. Wallingford, UK: CAB International. 2008. ISBN 0-85199-826-7.
- Taylor EL, Taylor TN. The Biology and Evolution of Fossil Plants. Englewood Cliffs, New Jersey: Prentice Hall. 1993. ISBN 0-13-651589-4.
延伸阅读
[在维基数据编辑]
外部連結
| 查看维基词典中的词条「真菌」。 |
| 關於Fungus 的圖書館資源 |
- Tree of Life web project: Fungi (页面存档备份,存于互联网档案馆)
- Mushroom Observer (mushroomobserver.org (页面存档备份,存于互联网档案馆)), a collaborative fungus recording and identification project
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||
|
|