
| 哺乳动物 化石时期:
| |
|---|---|
  | |
|
科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 总纲: | 四足总纲 Tetrapoda |
| 演化支: | 爬行形类 Reptiliomorpha |
| 演化支: | 羊膜動物 Amniota |
| 演化支: | 合弓類 Synapsida |
| 演化支: | 哺乳型类 Mammaliaformes |
| 纲: |
哺乳纲 Mammalia Linnaeus, 1758 |
| 现生子类 | |
哺乳动物或哺乳类是对脊椎动物下哺乳纲(学名:Mammalia)动物的统称,现存的物种主要分属胎盘类(绝大多数)、有袋类和单孔类三个演化支。与同属羊膜动物、在石炭紀末期(約3億年前)分化的爬行類、鳥類不同的是,哺乳类具有新皮质、毛皮、三個听小骨和乳腺,雌性还會用乳腺分泌乳汁餵養幼崽。目前,已得到人類正式記錄的哺乳類物种约有6400个,其中物种数量最多的目有囓齒目、翼手目和真盲缺目(刺猬、鼹鼠、鼩鼱等),多樣性略低於以上三目的有灵长目(包含人類在內的猿、猴等)、偶蹄目(鯨豚類和其他偶蹄目)和食肉目(猫、狗、海豹等)。
以支序學的角度來看,哺乳類是合弓綱中的唯一成員,而合弓綱和蜥形綱組合成了一個更大的演化支羊膜動物。哺乳動物的先祖為盤龍目中的楔齒龍類。在約3億年前,亦即石炭紀末期,合弓綱和蜥形綱從羊膜動物中分化出來後,蜥形綱則成為了現生的鳥類和爬行類。在獸孔目出現之前的二疊紀早期,楔齒龍類的基群演化成了好幾支非哺乳類合弓綱,這些生物偶爾會被誤認為類似哺乳類的爬行類。現生的哺乳類起源於非鳥恐龍滅絕之後,約6600萬年前的新生代,自出現以來便是陸生動物中最具優勢的類群。
哺乳類的基本体型为四足步行型,大多数哺乳动物可以使用其四肢在陆地上移动。同时,也有一些生活在海裡、空中、树上、地下等环境的哺乳动物,靠着經由演化產生的四肢末端形态,來适应不同的生活环境。哺乳动物小至30至40毫米大的凹臉蝠,大到30公尺長的蓝鲸(是地球史上最大动物)。其寿命在2年(鼩鼱)到211年(弓頭鯨)之间不等。除单孔目之下的产蛋物种外,其他现存哺乳动物都会在分娩时直接诞下幼崽。哺乳动物中最大的一群动物是被称为胎盤動物的分类阶元,它们的胎盤可以在孕期为胎儿供养。
绝大多数哺乳动物具有认知能力,有的还有大容量的脑和自我意识,会使用工具。哺乳动物可用多种不同的方式发声、交流,如发出超聲波、用气味标记领地、发出警告信号、鸣唱、回声定位等。哺乳动物会组织形成裂变融合社会、一雄多雌制度和等级制度——但也有的更独来独往,有着自己的领地。大部分哺乳动物实行“一夫多妻制”,但也有的是“一夫一妻制”或者“一妻多夫制”。
在新石器革命期间,人类驯化了多种哺乳动物,导致农业活动取代狩獵採集,成为人类的主要食物来源,社会结构亦由游獵转向定居,各部落之间开始互相合作,发展出了人类的首个文明。自那时起,驯化的家养哺乳动物一直劳作在人类社会的交通与农耕上,它们向人类社会输出了食物(如肉类和奶制品)、毛皮和皮革。人们还会狩獵哺乳动物、将其用于体育竞技中,或是将其作为模式生物进行研究。自旧石器时代開始,哺乳动物即广泛在各种艺术创作中亮相,在文学作品、电影、神话、宗教中频频现身。然而,人类对哺乳动物栖息地的破坏(以森林砍伐为主)和偷獵活动导致了多种哺乳动物数量锐减乃至灭绝。
分类

自从卡尔·林奈首次将哺乳类分为一纲以来,哺乳动物的定义曾多次变动。目前,人们对哺乳动物的分类方式还没有明确的共识;最新有关方面的研究由麦肯纳与贝尔(1997年)和威尔森与里德尔(2005年)作出。喬治·蓋洛德·辛普森的“哺乳动物分类与分类原则”(AMNH Bulletin v. 85, 1945)从系統分類學上阐述哺乳动物的起源和关系,他的理论直到20世纪末都还出现在各个教科书中。自辛普森发明分类系统以来,化石记录已几经修正。分类系统理论本身也经多方讨论、发展壮大,在支序分類學出现后更是愈演愈烈。虽然实地考察中新发现的生物证据让辛普森的分类系统逐渐落伍,但他的分类方式依然是哺乳动物分类体系中最为权威的。
现存哺乳动物中物种最多的六个目,属于胎盘动物。其中,物种最多的三个目是啮齿目(鼠、豪豬、河狸、水豚等哺乳动物)、翼手目(蝙蝠)和真盲缺目(鼩鼱、鼹鼠、溝齒鼩等)。接下来的三大目取决于选择什么分类方法,可以是灵长目(如猿、猴和狐猴)、鲸偶蹄目(如鯨和偶蹄目)和食肉目(如猫、狗、 鼬、熊、海豹等)。2005年,第三版的《世界哺乳动物物种》共列出了5416个物种,分为1229属、153科、29目。2008年,國際自然保護聯盟(简称IUCN)在整理其瀕危物種紅色名錄时完成了一项历时五年的全球哺乳动物评估计划,共收入了5488个物种。2018年发布在《哺乳动物学杂志》的研究指出,现已知哺乳动物物种共6495个,其中包括96个最近灭绝的物种。
定义
“哺乳动物”是一个现代生物学概念,其学名Mammalia由卡尔·林奈在1758年提出,出自拉丁语當中有“乳头、流食”之意的mamma。在一篇1988年的文献中,蒂姆·罗(Timothy Rowe)从系统发生学角度将哺乳类定义为哺乳动物的冠群,使之成为现生单孔目(针鼹和鸭嘴兽)、獸亞綱(有袋類和胎盤動物)的最近共同祖先所在的演化支,该演化支中含有该祖先的全部后代。鉴于这个祖先生活在侏罗纪,罗的定义不包含任何三叠纪早期的动物,哪怕19世纪中期就已经依化石将三叠纪贼兽目划为哺乳动物。如果将哺乳动物认为是一个冠群,那么它的起源的时间可以由与现生哺乳动物亲缘更接近的动物的出现来估计。比如说,与兽亚纲相比,昂邦兽与单孔目更接近;环齿兽属与双兽属则与兽亚纲更接近。上述三个属的化石在1.67亿年前的侏罗纪中期都有发现,所以这应该是对这一冠群出现时间的合理的估计。
湯姆·坎普(Tom S. Kemp)曾提出过更为传统的定义:“拥有下颌骨–鳞状骨颌关节的合弓纲动物,上下臼齿应能咬合,咬合过程中牙齿应能横向移动”。坎普还提出另一定义,即当前存活的哺乳动物与中國尖齒獸的最近共同祖先所形成的演化支。在他看来,这两个定义互相等价。目前发现的满足坎普定义的动物中,最古老的当属2.25亿年前出现的提基兽,因此从这一角度而言,哺乳动物早在三叠纪晚期就已出现。
麦肯纳/贝尔分类
1997年,马尔科姆·C·麦肯纳和苏珊·K·贝尔(Susan K. Bell)将哺乳动物整个重新分类,其成果后来被称作麦肯纳/贝尔分类。1997年,二人著书《对物种以上级别的哺乳动物种群的分类》(Classification of Mammals above the Species Level)研究了史上所有哺乳动物属及以上的生物分类,对它们的系统发生、关系、分布进行了描述,但有几个高阶分类与后来的分子基因数据不符。二人都是美国纽约自然史博物館的古生物学家。麦肯纳从辛普森那里得到灵感,与贝尔一道对全体现生、已灭绝哺乳动物进行分类,反映了历史系谱学观点。
在以下列表中,已滅絕的演化支由劍標(†)標註。分类阶元地位依照麦肯纳与贝尔的观点,所使用名称与现名不同。
哺乳綱 Mammalia
- 原兽亚纲 Prototheria
- 獸形亞綱 Theriiformes:現存的哺乳動物與其已滅絕的近親
胎盘动物的分子生物学分类
在21世纪早期,在经过对DNA分析后,分子生物学家对哺乳动物科群关系有了新的认识。这些新发现大部分已被反转录转座子的出現-未出現資料独立证实。基于分子研究的分类系统将胎盘动物分为三大类/谱系——非洲獸總目、异关节总目和北方真獸高目,分子钟推定在白垩纪时期发生分化。这三个谱系之间的关系尚有争议,目前人们通过假定不同的基群,共提出过三个假说——以北方真兽高目为基群的大西洋獸類、以异关节总目为基群的上兽总目和以非洲獸總目为基群的外非洲胎盘类。北方真獸高目下含两个主要分支——靈長總目和勞亞獸總目。
这三个胎盘类谱系据估计在1.2亿至1.05亿年前分化,具体分化年代取决于估算时用的是哪种DNA(如核DNA、线粒体DNA)、对古地理学数据的阐释方式等。
| 哺乳綱 Mammalia |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

本分支图係基於詹姆士·塔沃(James E. Tarver)等人在2016年的研究。
第1類群:非洲獸總目 Afrotheria
第2類群:异关节总目 Xenarthra
第3類群:北方真獸高目 Boreoeutheria
演化
起源
包含了現存的哺乳綱以及其他已滅絕近親的合弓綱最早起源於約3.23亿年前至3億年前的賓夕法尼亞世,與其他爬行類及鳥類的演化支分歧。哺乳綱的冠群演化自侏儸紀早期的哺乳形类物種。底下為以哺乳形類做為冠群的分支圖。
| 哺乳形类 Mammaliaformes |
|
||||||||||||||||||||||||||||||||||||||||||||||||
从羊膜动物中分化

最早的陸生脊椎动物為羊膜動物,和其他四足類一樣,牠們也具有肺臟及四肢。然而羊膜動物的卵內層有膜,可將水分鎖於其中,因此得以在乾燥的陸地上產卵、正常发育胚胎;相較之下,兩棲類僅能將卵產於水中。
最早的羊膜動物出現於石炭纪的賓夕法尼亞世,由早期的爬行形类演化而來,而當時在陸地上已有許多昆蟲及其他無脊椎動物棲息,陸棲的植物則包括链束植物、藓类植物门等。数百萬年後,羊膜動物分歧為兩個演化支:合弓綱,包含了所有哺乳動物的共同祖先;以及蜥形纲,包含了龜、蜥蜴、蛇、鱷、恐龙與鳥類。合弓綱在顱骨的兩側後方僅具有單一開孔,稱為顳顬孔。合弓綱下的盤龍目包括了二叠纪早期最大型與最兇悍的物種。非哺乳類的合弓綱物種過去曾被統稱為「似哺乳爬行動物」。
合弓綱中的獸孔目,由盤龍目的楔齿龙类演化而來,出現於二疊紀中期(約2.65億年前),為當時最為興盛的陸生脊椎動物。牠們在顱骨及下顎上有些許特徵與真盤龍亞目基群物種相異。最早的獸孔目物種外型與楔齿龙类十分類似,而晚期的獸孔目物種如犬齒獸亞目的新頜獸小目中,有些外形已經與哺乳動物無異。從獸孔目到哺乳動物的演化依據下列特徵分為多個階段:
- 第二骨質硬颚的漸進式發展
- 四肢逐漸由身體兩側移轉至下側,使動物於移動時能避免卡里爾約束進而提高運動的持久度。但這項特徵的演化過程十分緩慢且非常不穩定,比方來說,所有草食性的非哺乳類形獸孔目物種四肢仍為兩側延展(部分較晚期物種則為半直立)、二疊紀時期的肉食性獸孔目物種期前肢為兩側延展、較晚期的肉食性物種則具有半直立的後肢。而現存的單孔目則仍然保留半直立式的四肢。
- 下颌骨逐漸成為下顎主要的骨骼組成,在三疊紀時,哺乳類的下顎僅包含有下颌骨,而其他原本組成下顎的骨頭則後移形成中耳。
哺乳动物祖先
2.52亿年前,地球上出現了多宗滅絕事件,它們所引起的連鎖效應令大多中大型獸孔目物種滅絕。三疊紀早期時,中型與大型的陸生肉食生態位逐漸被主龙形类取代。少數大型犬齒獸亞目物種如犬齒獸屬與橫齒獸科的物種仍分別保有大型肉食與草食性動物的生態位,但到了侏儸紀時,所有大型陸生草食動物的生態位均被恐龍所取代。
最早的哺乳動物(克坎普的定義下)出現於約2.25亿年前的三疊紀晚期,在最早的獸孔目出現4000萬年之後。這些哺乳動物於侏儸紀中期時保有夜行性食虫动物的生態位,但也逐漸有多樣化的發展。例如,生存於侏儸紀時期的獺形狸尾獸,具有與現存水獺類似的外型,擅長游泳、挖掘及捕捉魚類。然而,當時幾乎所有的哺乳動物均為夜行性,稱為夜行瓶頸,許多夜行性特有的特徵於現存的哺乳動物(縱使為晝行性)上仍然可觀察到。生存於中生代的哺乳動物主要為多瘤齒獸目、真三尖齒獸目與鼴獸類。最早已知的後獸下綱為中國袋獸,發現於中國東北辽宁省地層年代1.25亿年前白堊紀早期的页岩中。該化石十分完整,甚至保留了毛簇以及軟組織的印痕。

目前已知最早的真獸下綱為外型類似鼩鼱的中華侏羅獸,生存於1.6亿年前的侏儸紀晚期。更為晚期出現的始祖獸,出現於距今1.25亿年前的白堊紀早期,具有許多有袋類共有而有胎盤類缺乏的特徵;這代表著有部分有袋類的特徵曾經存在於有袋類與有胎盤類的共同祖先上,而這些特徵在之後有胎盤類的演化支中逐漸退化或喪失。例如,由骨盆向前延伸的上恥骨並不存在於現存的有胎盤類物種上,但可於有袋類、單孔目、非獸亞綱哺乳動物、多瘤齒獸目以及烏克哈獸(生存於白堊紀早期的真獸亞綱亞洲掘蝟目物種)上發現。上恥骨主要用於在運動時能支持腹部,但這也限制了妊娠期胎兒發育的空間。狹窄的發育空間代表胎兒出生時的體型必須非常小,也因此不得有太長的妊娠期,為現存有袋類的共有特徵。而胎盤的演化可能發生於上恥骨的退化消失之後。
目前已知最早的單孔目物種為泰諾脊齒獸,生存於距今1.2亿年前的澳洲。單孔目物種仍然保留有許多早期羊膜動物的特徵,包括用於排泄、排遺、生產的單一開孔(泄殖腔),以及革質未鈣化的卵,這些特徵在現存的爬行類或鳥類身上可以發現。
哺乳動物特徵的出現
生存於距今1.95亿年前侏儸紀早期的吴氏巨颅兽,為最早發現下顎僅由鱗骨與齒骨組成的物種,早期合弓綱常見的關節骨已經退化。

最早明確的毛髮化石證據來自於距今1.64亿年前侏儸紀中期的獺形狸尾獸與哺乳形巨齒獸。1950年代,位於犬齒獸亞目物種上颌骨與前上頜骨上的小孔被認為是其感覺毛血管及神經的通道,是為間接毛髮存在的證據;然而,這些小孔其實並無法證明感覺毛的存在,例如現存的雙領蜥其上颌骨上也具有和三尖叉齒獸類似的小孔,但是並不具有感覺毛。不過現今大多的復原圖仍然會將三尖叉齒獸加上感覺毛。針對二疊紀粪化石研究顯示,當時的非哺乳類合弓綱已經具有毛髮,這代表毛髮的存在可以向前推至二齒獸下目。
目前仍無法得知哺乳動物是在何時演化為內溫動物,但多半認為獸孔目是哺乳動物演化支上最早出現的內溫動物。現存的單孔目物種比起有袋類與有胎盤類具有較低的體溫與變化率較大的代謝速率,然而有證據顯示有些較古老的單孔目物種,包含獸亞綱的祖先,可能具有與現存獸亞綱相近的體溫。此外,獸亞綱中非洲獸總目與異關節總目部分現存的物種具有為了適應環境重新演化出的低體溫。
哺乳動物直立式四肢的演化並不完全,現存與已滅絕的單孔目四肢均為兩側延展。向下延展的四肢特徵最早約出現於侏儸紀晚期至侏儸紀早期,生存於距今1.25億年前的始祖獸與中國袋獸均具有向下延展的四肢。上恥骨最早發現於三瘤齒獸科,為牠們與哺乳形類的共有衍徵,於現存的非有胎盤類哺乳動物身上仍可發現。然而於大帶齒獸與紅層獸中上恥骨已經退化消失。
一般認為乳汁的產生最早是為了讓產下的卵保持潮濕,之後才發展出哺乳的行為。這樣的論據主要來自於對現存單孔目(卵生哺乳動物)的觀察。
哺乳动物的崛起
獸亞綱哺乳動物於距今6600萬年前白垩纪﹣古近纪灭绝事件發生後的新生代開始填補原先由非鳥恐龍、其他蜥形綱及其他合弓綱所遺留下來的中大型生物生態位空缺,並且體型上有十分顯著的成長(巨型動物群)。在滅絕事件發生後,尚未滅絕的鳥類與哺乳類展現了十分快速的輻射演化。例如,目前已知最古老的蝙蝠,爪蝠與伊神蝠,發現於約5200萬年前,在滅絕事件發生後僅經過了1400萬年,然而外型已接近現存的蝙蝠並具備了完整的飛行能力。
分子類緣關係之研究原先認為大多胎盤類下的目約於1億年至8500萬年前分家,而現存大部分的科則於始新世至中新世期間出現。然而,目前並沒有發現生存於白堊紀的胎盤類哺乳動物化石,已知最早的胎盤類哺乳動物出現於非鳥恐龍滅絕之後,過去認為是古新世早期的古蹄獸,然而近年來的研究將牠們重新分類為非胎盤類的真獸下綱物種。在基因與型態變化率的重新校準之後,普遍認為胎盤類哺乳動物起源於马斯特里赫特期,而現存大多的演化支則起源於古新世。
目前已知最古老的靈長目物種為距今5500萬年前的阿喀琉斯基猴,體重僅有20至30公克,與人類的手掌大小相當。
解剖学结构
特征
现生的哺乳动物都有汗腺,某些物種的汗腺分化为可以产奶的器官以哺育幼崽。然而,在分類化石的時候,汗腺並沒有辦法作為辨別哺乳動物的特徵,因為化石無法保存軟組織。
早期哺乳动物具有许多共同特征:
- 顎关节——下颌骨和鳞骨(颅骨中的一块小型骨骼)连接,形成下颚。大多数有颔下门动物(包括早期的獸孔目)的颚关节还有关节骨(下颚后方的小骨)和方骨(上颚后方的小骨)。
- 中耳——在冠群哺乳动物中,声音从鼓膜通过三块连接在一起的骨骼传入耳内,这三块骨骼分别是锤骨、砧骨和镫骨。锤骨和砧骨由关节骨和早期兽孔目下颌关节的方骨演化而来。
- 换牙——有些物種终生只會換一次牙(再出齿),有些則完全不换牙(一出齿),如齒鯨和鼠科。大象、海牛和袋鼠終生會不斷地換牙(多出齿)。
- 釉柱釉质——牙齿表面的牙釉質由棱柱状固体结构构成,从牙本質一直延伸到牙齿表面。
- 雙枕髁——哺乳動物的枕骨後頂有兩個關節髁,與寰椎相接;然而多數四足類僅具有單枕髁。
在哺乳动物的三叠纪祖先身上,这些特征基本不存在。除了现代胎盘动物之外,几乎所有的哺乳型动物都有上耻骨。
两性异形
平均來說哺乳類雄性體型會大於雌性,在已被學者研究的物種中,超過45%的物種雄性體型至少比雌性大10%。大多數哺乳類的兩性異形傾向於雄性,有些則不具有兩性異形,有些則傾向於雌性(如兔形目)。物種體型越大,兩性體型的差異越大(倫施法則),這很可能是因為雄性與雌性在體型上分別承受了不同的選汰壓力。傾向於雄性的兩性異形則與雄性性擇有關,雄性互相競爭以爭取交配權,性擇壓力越大,雄性的體型差異就越大。在其他哺乳類中,性擇壓力也與雄性與雌性的體型大小有關。至於雌性的選汰壓力,學者發現一夫多妻程度越高的物種,斷奶的時間越晚,就算去除掉體重差異,校正潛在的偏差後,結果亦然,除此之外,體型較大的雌性繁殖速度較慢,顯示出對於體型較小的雌性來說,生殖選擇的壓力較大。
上述內容適用於大部分的哺乳類,但須注意的是不同的目之間還是會有差異。
生物系统
大部份的哺乳動物有七塊頸椎骨, 譬如蝙蝠、長頸鹿、鯨魚和人類。海牛屬和二/三趾樹懶屬都是例外,前兩者只有六塊,後者則有九塊。所有哺乳動物均有新皮質,僅存在於哺乳動物的腦部位。與單孔目及有袋類不同,胎盤動物有胼胝體。
哺乳動物的肺部呈海綿狀及蜂巢狀。呼吸主要通过橫膈膜进行,该结构将胸腔、腹腔一分为二,呈向胸腔凸入的形状。横膈膜收缩时形状变平,增加肺的容积。空气通过口腔、鼻腔进入身体,再流经喉、气管、支氣管,最后进到肺泡里,将肺泡张开。横膈膜舒张则会有相反的效果,降低肺容量,将空气压出体外。在运动时,腹壁收缩,增加横膈膜的压力,可使空气的进出频率增加、力度增大。肋骨架也可通过其他呼吸系统肌肉扩张、收缩胸腔,让呼吸的空气永远向气压低的方向移动。这种肺因结构类似铁匠的风箱而被称作“风箱肺”(bellows lung) 。 哺乳动物的心脏有四个腔室,上面两个是心房,负责接受血液;下面两个是心室,负责泵出血液。心脏有四个瓣膜,能将各个腔室分开,确保血液沿正确的方向流动(防止回流)。肺毛细血管中的血液在氣體交換结束后会变为富氧血,经四根肺静脉源源不断地汇入左心房,随后再经开口流入左心室。心房、心室舒张时,大部分血液会依照既有方向流动,但在心室舒张末期时,左心房会收缩,将血液打入心室中。和其他肌肉类似,心脏需要靠血液中的营养物质、氧气才能正常工作,这些养分由冠状动脉血提供。


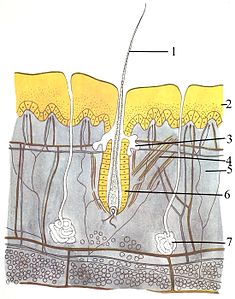
哺乳动物的外皮系统由三层皮肤组成——表皮、真皮和皮下組織。表皮通常有10至30个细胞厚,主要作用是防水。表皮层最外层的细胞不断脱落,而最里层的细胞会不断分裂、将现有的细胞组织向外推。再往深处的真皮层是表皮层的15至40倍厚。真皮层由血管、骨性结构等许多部分构成。皮下组织由脂肪組織构成,能储存脂质、提供缓冲和隔热功能。皮下组织的厚度在各个物种之间差异极大。海洋哺乳動物为了保暖,长出了厚厚的皮下组织(鯨脂),其中露脊鯨的鯨脂最厚,可达51公分。部分其他非哺乳动物长有触须、羽毛、刚毛、纤毛等类似毛发的结构,但只有哺乳动物身上才生有真正的毛发,是哺乳动物类的一大决定性特征。有的哺乳动物的毛发可能不明显,但经过仔细检查总能发现毛发痕迹,通常是位于身体较为遮掩的部位。


食植动物的身体结构多种多样,以便进食、消化植物。哺乳动物的牙齿演化出的结构可以将完整的植物组织切开。比如说,食果动物和食用软叶子动物有低冠齿,可以磨碎叶子和种子。食草动物喜食坚硬、硅含量高的草,它们的高冠牙能够磨碎坚硬的植物组织,比低冠齿更耐磨。大部分肉食哺乳动物长有裂肉齒(齿的长度因饮食结构而有区别)和长长的犬齿,其换牙模式也较为接近。
偶蹄目的胃分为四个部分:瘤胃、蜂巢胃、重瓣胃和皺胃(只有反芻动物有瘤胃)。瘤胃和蜂巢胃负责将摄入的植物组织与口水搅拌,分为固体和液体。随后,固体部分被压成食团,再度返回口中,让舌头挤压出汁液并吞咽回去。攝入的食物进入瘤胃和蜂巢胃后,分解纖維素的微生物(细菌、原生生物和真菌)会產生分解植物中的纤维素所需要的纤维素酶。與反芻動物相反,奇蹄目會將離開胃的已消化食物儲藏于擴大的盲肠中讓細菌發酵。 食草動物複雜的消化系統對於分解粗糙又堅硬的植物纖維來説是必需的,相比之下,食肉動物只有一個結構簡單的胃,適應于主要消化肉類。食肉動物的盲肠不是已經消失就是結構簡單而短小,而大腸并沒有具備囊狀組織或者寬過小腸太多。

哺乳動物的排泄系統包括許多組成部分。與大多數陸生動物相似,哺乳動物是排尿素型動物,通過肝臟將氨轉化爲尿素實現尿素循环。而胆红素作爲血细胞中生成的副產品,會在肝臟生成的酶的幫助下,通過胆汁和尿排出體外。膽紅素藉助膽汁經過消化道的過程使得哺乳動物的糞便呈現出獨特的棕色。哺乳動物腎臟的典型特徵包括腎盂和腎錐體,以及由於細長的亨利氏環的存在而可以明確區分開的肾皮质和腎髓質。只有哺乳動物的腎臟呈豆狀,但也有少數例外,如鰭足類、鯨豚類和熊類的多頁腎臟。大部分成年胎盤動物體内都沒有泄殖腔的殘留痕跡。在胚胎時期,胚胎泄殖腔會發育成爲前部和後部,後部成爲肛門的一部分,而前部的發育則取決於個體的性別:雌性會發育成爲包括尿道和阴道的前庭,而雄性則發育成爲陰莖尿道。 不過马岛猬科、金毛鼹科和部分鼩鼱科成年後仍會保留泄殖腔。在有袋類中,生殖道與肛門雖然是分開的,但是外部仍然留有原始泄殖腔存在的痕跡。單孔目的学名“Monotremata”出自古希臘語的 μονός(monós,单一的)与 τρῆμα(trêma,孔),它們擁有真正的泄殖腔。
发声结构

與其他四足類一樣,哺乳動物有一個可以迅速打開和關閉的喉以發出聲音,以及一個能過濾這種聲音的上喉聲道。肺和周圍的肌肉組織可以提供發聲態所需要的氣流和壓力。而喉部控制著聲音的音高和響度,不過肺部呼出時的力量也有助於響度的提高。更原始的哺乳動物,例如針鼹,只能發出嘶嘶聲,因爲聲音是通過部分封閉的喉嚨呼出實現的。剩下的哺乳动物用声带发出声音,而非像鸟类的鸣管。聲帶在震动和绷紧时可以產生多種不同的聲音,例如呼嚕聲和尖叫聲。哺乳動物可以改變他們喉部的位置,在吞嚥時可以繼續通過鼻子呼吸,并且可以同時發出口腔音和鼻音,例如:狗的嗚嗚聲一般來説都很柔和,這是鼻音;而狗的叫聲一般都很響亮,這是口腔的聲音。
部分哺乳動物擁有較大的喉嚨,因此音高較低,例如錘頭果蝠的喉部可以占據整個胸腔並把肺、心臟和氣管推入腹部。大的聲帶也可以降低音高,如大猫的低吼。一些哺乳動物還可以發出次声波,如非洲象和鬚鯨。喉部較小的小型哺乳動物可以發出超聲波,可被特殊发育出的中耳和耳蜗探測到。在中生代时期,鸟类和爬行類是哺乳类最大的天敌,但鳥類和爬行類聽不見超聲波,那段时期超声波对哺乳类至关重要。一些啮齿类动物的母親和幼崽会用這一秘密的通信方法交流,蝙蝠也能用超声波回聲定位。齒鯨也會使用回聲定位,不過,與使用聲帶上延伸出來的聲膜所不同,齒鯨是通過額隆來控制聲音的。一些哺乳動物(如靈長類)的聲帶上附有一個氣囊,可以用來降低共鳴或者增大音量。
哺乳动物的发声系統由大腦中的腦神經核控制,其神经信号经由部分迷走神经(即喉返神經和上喉神經)传递。聲道则由舌下神经和顏面神經传递信号。對哺乳動物中腦内的导水管周围灰质區域进行電刺激會引起發聲。哺乳动物中,只有人類、海豹、鯨類和大象(可能也有蝙蝠)能够学习发出新声音。人类之所以有这一能力,是因为控制動作的運動皮層和脊髓中的运动神经元直接相連。
视觉
除了部分灵长目和后兽下纲动物外,其他所有哺乳类都是严格的二色视觉者,但其上级分类脊椎动物大都是四色视觉者。这是因为在演化早期为了避免与占据生态位优势的主龙形类在同一时间活动,哺乳动物大多是夜行性的,于是在这段漫长的时间里便逐渐丧失视网膜中四种视锥细胞中的两种。然而因祸得福,哺乳动物的夜视能力比大多数其他脊椎动物要强。部分灵长目的祖先通过基因突变获得了三色视觉。
皮毛

哺乳動物皮毛的主要功能為體溫調節,其他功能則包括保护、感測環境、防水與偽裝,不同形式的皮毛具有不同的稱呼及功能:
- 定期毛髮(Definitive)– 當達到一定長度後即會脫落。
- 感覺毛(Vibrissae)– 用於感測周遭環境。
- 毛皮(Pelage)– 包括護毛、絨毛與芒毛。
- 棘刺(Spine)– 堅硬的毛髮,用於防衛(例如豪豬背上的長刺)。
- 剛毛(Bristle)– 主要用於展示視覺訊號(例如獅子的鬃毛)。
- 毳毛(Velli)– 用於保暖,主要常見於新生的哺乳動物。
- 羊毛(Wool)– 長而柔軟的卷曲毛髮。
体温調節
毛髮長度與體溫調節無關:例如,部分棲息於熱帶地區的哺乳動物,如樹懶,它们的毛发长度和极地哺乳動物的毛发长度几乎相等,但並不具有与之对应的保暖能力;反而,部分僅具有短毛的熱帶哺乳動物,保暖能力卻與極區哺乳動物相當。毛髮生長的密度可以影響保暖的能力,尤其棲息於極區的哺乳動物的毛发极为浓密,例如麝牛除了具有30厘米長的護毛外,還具有高密度的絨毛,能有效鎖住並減少體熱的散失,讓牠們能於-40°C的環境下生存。有些沙漠哺乳動物,如駱駝,則透過高密度的毛來將外在環境的熱度隔絕於外,確保個體不會過熱;在夏天時,駱駝的毛髮溫度可達70°C,而皮膚溫度則能維持在40°C上下。水生哺乳動物的毛髮也十分浓密,可將水隔絕於外、保持皮膚乾燥,从而減少體熱散失。
体色

哺乳動物的體色源自於外在生存壓力,主要為偽裝、性選擇、溝通、及體溫調節。皮膚與毛髮的顏色主要由黑色素的種類與數量決定。真黑素構成了棕色及黑色;棕黑色素則構成了黃色及紅棕色,使個體帶有大地色調。有些哺乳動物,如山魈,由於结构色具有更為鮮艷的體色。許多樹懶為綠色,因為牠們的毛髮中带有藻類,這些藻類與樹懶共生,可帮助树懒偽裝。
大多哺乳動物都會仰賴牠們的體色來進行偽裝,減少被獵物或掠食者發現的機會。棲息於北極及亞北極地區的哺乳動物,如北極狐、環頸旅鼠 、白鼬、白靴兔,夏天時體色是棕色,而在冬天體色变為白色,這樣毛色的變化可帮助它们融入周遭環境。以靈長類和有袋類为主的樹棲哺乳動物在身上的某些部位具有紫色、綠色或藍色的斑塊,這樣的趋同演化顯示這些斑塊可能有助於牠們在樹棲的環境中生存。
部分具有自我防衛能力的哺乳動物,例如具有強烈氣味的臭鼬、以及兇悍的蜜獾,具有黑白相間的毛皮,能發揮警戒作用。 哺乳動物的體色有時為兩性異形,常見於靈長目物種。雌性與雄性間的體色差異可能代表了攝取營養量或是荷爾蒙強度不同,而這些通常是選擇配偶的重要因素。體色也會影響到身體維持熱量的能力,較深色的毛髮可以幫助哺乳動物從太陽輻射中吸收較多的熱量。部分小型哺乳動物,例如田鼠,在冬天時具有較深色的毛髮以用於保暖。另外,斑馬身上黑白相間的條紋似乎可以降低牠們被蚊蟲侵擾的機率。
生殖系统

雄性胎盘哺乳动物的阴茎兼具排尿和交配功能。不同物种勃起的方式不同,有的是靠海绵体血管充血,有的是用肌肉运动。有的阴茎在疲软时会收入阴茎鞘中,此外有的胎盘动物还生有陰莖骨。有袋类动物的阴茎通常会分叉,针鼹的阴茎则通常有四个末端,其中只有两个能正常工作。绝大多数哺乳动物的睾丸位于阴囊中,通常这一结构位于阴茎之后,但有袋目的阴囊在阴茎之前。雌性哺乳动物通常拥有陰蒂、大陰唇、小陰唇等外阴结构,体内则通常有成对的输卵管、一至两个子宫、一至两个子宮頸以及一条阴道。有袋目由两条侧阴道和一条中阴道。单孔目动物的“阴道”结构类似于“泌尿生殖窦”。胎盘动物的子宫形态各异,有的是双子宫、双宫颈共享一条阴道的重複子宮;有的是两个子宫角接入一个宫颈、一条阴道的對分子宮;有的是两个子宫角在远端相连,又在中线处分开,呈“Y”形状,称之为雙角子宮;再有的只有一个子宫,称为單子宮。

哺乳动物的祖先繁殖时会娩出未完全发育的胎儿,要么是直接胎生,要么在出世前会暂时裹在软壳卵中。这一机制很可能是由于上耻骨结构导致胎儿躯干无法进一步生长。卡岩塔獸是已知采用这一繁衍方式的合弓纲中最古老的一属,分娩时诞下未发育的围产期幼崽,但大小比所有现代哺乳动物的幼崽都要大。绝大部分现存哺乳动物都是胎生,直接产下成型的幼崽。不过,单孔目其下的物种都是产蛋的。单孔目的性別決定系統和大部分其他哺乳动物的都不一样。其中,鸭嘴兽的性染色体不像兽亚纲的哺乳动物,倒与鸡的更为相像。
胎生哺乳动物均属于兽亚纲,其中现今存活的哺乳类分布在有袋类和胎盘类这两个下纲中。有袋类动物的妊娠时间较短,通常短于其发情期,未完全发育幼崽诞下后会在母亲体外继续发育。许多有袋目生有育幼袋,位于母亲的腹部。这是诸多胎生动物的共同祖征;非胎盘类动物的下耻骨导致胎儿躯干分娩前无法完全发育。就连不属于胎盘类的真兽下纲都是如此繁衍的。胎盘类哺乳动物的幼崽出生时相对发育较为完善,但母亲怀孕时间更长些。其名称“胎盘动物”中的“胎盤”二字指的就是连接胎儿、子宫壁之间的结构,可帮助胎儿获取营养。胎盘哺乳动物要么完全没有下耻骨,要么下耻骨已演变为阴蒂骨,好让胎儿躯干在母体内进一步发育。
哺乳动物经过特化的乳腺能够产奶,是幼崽的主要食物来源。单孔目动物由于分化出去的时间较早,没有绝大多数哺乳动都有的乳頭,但却有乳腺结构,幼崽会舔舐母亲腹部的哺乳斑块来摄入母乳。和胎盘动物相比,有袋目动物因幼崽出生时发育不完善,其母乳不论是营养成分还是出奶频率都大不相同。此外,有袋目的乳腺各自独立工作,能在幼崽的各个发育阶段提供不同的母乳。乳糖為胎盤類母乳的主要糖份、而有袋類母乳的主要糖份則主要為寡醣。斷奶指的是哺乳動物在成長過程中,逐漸脫離母乳轉向攝食固體食物的過程。
温血状态
幾乎所有哺乳動物皆為内温動物,能透過毛髮留住熱量來維持體溫恆定,因此,比起爬行類與昆蟲等外溫動物,哺乳動物和鳥類可以在較為寒冷的環境中活動。内温動物透過攝取食物來提供熱量,因此需要的食物比體重相同的爬行類還多。小型食蟲哺乳動物食量驚人,例如普通鼩鼱每天就必須攝取相當於體重2至3倍的食物。少數物種幾乎不會產出代謝熱,因此被認為是變溫動物,如裸鼹鼠。另一個特例是樹懶,它们的體溫隨著環境溫度變化,範圍大約在25°C至35°C,最低時可能只有20°C,為異溫動物。鳥類也是內溫動物,因此內溫性並非哺乳動物專屬的特徵。
壽命
哺乳動物的壽命差異極大,如鼩鼱大約可以活2年,而已知最长寿的弓頭鯨則活到了211歲。影響壽命長短的因素目前未知,但數名學者認為主要的因素是DNA修復的能力。哈特(Hart)與塞特洛夫(Setlow)於1974年的研究顯示,在七種哺乳動物中,DNA切除修復的能力與壽命長短成正比。物種壽命不但與辨識DNA雙股結構斷裂的能力有关,也与DNA修復蛋白質Ku80的濃度有關。一個包含16種哺乳動物細胞的研究顯示,長壽物種負責修復DNA的基因較為活躍。一個包含十三種哺乳動物的研究顯示,細胞中DNA修復酶聚腺苷酸二磷酸核糖转移酶的濃度與壽命相關。其他研究也顯示數種哺乳動物的壽命與DNA修復能力相關。
运动
陆行

大多脊椎動物——包括兩棲類、爬行類與部分哺乳動物,均為蹠行,整個腳掌貼地而行;而部分哺乳動物,如貓或狗,則為趾行,行走時僅有部分腳趾著地,这样跨步距離更大,动物也因而能够以更快的速度進行奔跑。除了这些灵活性高的动作外,趾行的哺乳動物多半也具有較為安靜的移动方式。另外有部分哺乳動物,例如馬,為蹄行,行走時僅有趾尖上的角質蹄甲著地,進一步地提高跨步距離與速度。少數哺乳動物,如大型猿類、大食蟻獸與鴨嘴獸,前肢是以指背行走。部分哺乳動物以双足步行,例如人類;相較於四足著地的動物,双足動物擁有更好的視野、保存更多的體力,並可使用雙手操作物件、或是抓取食物。部分双足哺乳動物跳躍前行取代行走,例如袋鼠與更格盧鼠。
在不同地形、情境、移动速度下,動物會採用不同的步態。舉例來說,馬在自然情況下就具有四種步態,其中速度最慢的步態為常步,其他三種依據速度由慢至快分別為速步、驅步及襲步。動物有時也會有較為不常見的步態,例如側走或倒退。例如人類主要的步態為步行與跑步,但是在面臨其他環境時也會使用不同的步態,例如在狹窄的空間中,人類就會改用四肢匍匐的形式前進。哺乳動物的步態多种多样,在移動時附肢落下或舉起順序千变万化;根據這樣順序的排列,大體上能將步態分為數類。以四足動物為例,主要分為三類:行走步態、跑步步態以及飛躍步態。行走為最常見的步態,是指在任何時間均會有其中一足著地的狀態;而跑步則在某些時刻,會發生所有肢體均騰空不著地的情況。
飞行
蝙蝠是哺乳类中唯一真正可以飞行的。 它们通过上下扇动翅膀(通常还有一些前后的运动)可以在空中以恒定的速度飞行。蝙蝠本身的运动速度,与翅膀相对于空气的运动相叠加,可以在翅膀上形成更快的气流。这将会产生指向前方与上方的升力,与指向后方与上方的阻力。这些力向上的分量用于抵消蝙蝠自身的重力,使其保持在空中。这些力向前的分量是蝙蝠向前飞行的动力。
与鸟类相比,蝙蝠的翅膀更薄,有更多的骨骼,可更精细地控制翅膀,飞行时能提高的升力、减少阻力。蝙蝠在向上扇动翅膀时,可以将翅膀向身体一侧收起,这让它们在飞行中消耗的能量比鸟类减少了35%。它们的翼膜很容易受伤。不过翼膜上的组织可以再生,一些小的裂伤可以很快愈合。在蝙蝠翅膀的小突起上,有一些对触觉敏感的默克尔细胞。这种细胞也存在于人类的指尖。这些敏感区域的位置在不同种类的蝙蝠上是不同的。每个突起中心都有一根细毛,使得这些区域更加敏感,让蝙蝠可以检测和收集通过翅膀的气流信息。蝙蝠可以利用这些信息改变翅膀的形状,进一步提高飞行效率。
林栖

林棲動物大多擁有修長的四肢,可穿過縫隙取得果實或者其他資源、測試前方着力点的堅固程度,还可以在樹間擺蕩。像樹豪豬、侏食蚁兽、蜘蛛猴、袋貂般的林棲動物會卷起尾巴缠握樹枝。蜘蛛猴的尾巴尖端长有黏著墊或一小塊無毛的区域,可增大摩擦力。它们的爪子可在粗糙接觸面上幫助調整运动方向。松鼠爬树也是基于类似的原理——松鼠身体轻盈,其爪子提供的摩擦力可让它们在巨大的树干上如履平地。然而爪子也會干擾動物抓取細小的樹枝,因为在抓握时指爪可能会绕树枝一周,反过来刺入掌心。灵长类动物的指尖无毛,可在抓握时提供摩擦力,幫助動物將手固定在樹枝上。不過這種抓握方式會受摩擦力的角度影響,所以太過粗大的樹枝會導致抓握力下降。在下树时,一些林栖動物(例如松鼠)進化出了灵活性极强的踝關節,这使得它們的脚可以轉到完全“相反”的角度,好让爪子鈎住粗糙的樹皮以對抗重力。林棲動物的小體型帶給它們許多優勢,例如:增加了樹枝對它們的相對尺寸、降低了重心、增加了穩定性、減輕了重量(使得它們可以在細小的樹枝間穿梭)以及增加了在複雜的栖息地裏活動的能力。蜜袋鼯等滑翔動物的體型和體重对它们的滑翔能力影响极大。部分靈長類動物、蝙蝠,以及所有種類的树懒能够以各种角度将自己稳稳地挂在树枝上,只在失去抓力时才会跌落。
穴居与土棲


穴居哺乳動物(如獾、裸鼹鼠等)擅長挖掘洞穴,主要棲息於地底。許多啮齿目属于穴居哺乳動物,牠們大部分时间躲在自己挖的地穴中,只有少数时间會在地表活動。“土棲哺乳動物”完全在地底栖息,只演化出一部分适应地底活动的则被称为“半穴居哺乳动物”。有些穴居物種会钻入地下來调节体温、躲避掠食者的捕食和儲藏食物。
穴居哺乳動物多半具有梭形的身體,在肩膀處最寬,尾巴及口鼻部兩端收窄。由於长时间置身于不可見光的地穴中,許多穴居哺乳動物的眼睛均有退化的現象,不同物種的退化程度不同。大多半穴居哺乳動物的眼睛仍未退化;囊鼠的眼睛雖然非常小,但仍然保有作用。不过,完全穴居的哺乳動物(如袋鼴)的眼睛已經完全退化,比如鼴鼠的眼睛退化為痕跡器官,金毛鼴的眼睛表層更是直接被皮膚覆蓋。和眼睛类似,穴居哺乳動物的外耳廓也大多偏小或退化。完全穴居的哺乳動物大多擁有較粗短的四肢,可爆發出更強的力量;而許多半穴居哺乳動物則仍然保有适合奔跑的四肢。穴居动物的前掌大多寬厚,能在挖掘地穴時將土挖鬆;後掌則大多具有蹼和爪的结构,能將這些挖鬆的土向後推。部分穴居哺乳動物的門齒较大,可防止泥土灌入口內。
鼩鼱、刺蝟及鼴鼠等穴居哺乳動物最早被分類至食虫目,但這個目別現在已被廢棄。
水生
鯨豚與海牛等完全水生的哺乳動物,它们的後肢已經演化成尾鰭來在水中推進。鯨豚透過上下扭動身體來帶動尾鰭於水中高速推進,特化為胸鰭的前肢則能操控方向。許多动物还有背鰭,可防止牠們在游泳時腹背翻轉。海牛同樣透過上下擺動尾鰭來於水中推進,並能透過傾斜來轉向,前肢則如槳一樣能協助轉彎或是做為剎車減緩推進的速度。
半水棲的哺乳動物(如鰭足類)同時具有特化如槳的前後肢,肘關節與踝關節则埋藏於軀幹內。这些动物的身体经过演化,可以将水的阻力尽量减少。它们拥有流线型的身体,皮肤中的肌束还能增加身体四周的层流,让它们轻松掠过水中。另外牠們也没有立毛肌,好让毛髮在游泳時能維持平鋪,確保身體保持流線形。牠們主要透過拍動槳狀的前肢來推進,類似於企鵝與海龟;桨状前肢的运动并不是连贯的,动物在每次划水之间用滑动体态保持前行。和陸生的食肉目物種相比,鰭足類具有較短的前肢,形成較短的力矩讓牠們附著於肩膀與肘關節的肌肉能夠爆發出更強的力量。後肢部分則主要用來穩定游泳時行進的方向。其他半水棲的哺乳動物則包括河狸、河马、水獺及鴨嘴獸。河馬為十分大型的半水棲動物,长有粗壯的骨骼,可支撐起它们龐大的體重,同時較高的比重讓牠們得以沉至水底並於河床上行走。
行为
沟通与发声

许多哺乳动物通过发出声音来交流。声音交流可以达成诸多目的,比如求偶、警戒、指明食物来源、社交等。雄性在求偶时发出的声音可屏退其他雄性、吸引雌性,比如狮子和红鹿的吼声。座头鲸的鲸歌可以吸引雌性;它们在不同海域时,发出的声音也会不一样。某些动物的叫声具有社交功能,如长臂猿的领域叫声。长矛鼻蝙蝠还能用不同的声音频率来分辨不同的群体。长尾黑颚猴能发出不同的警示音来针对至少四種捕食者,而猴群也會依據警示音的不同做出不同反應。舉例來說,如果警示音警示有蟒蛇出現,則猴群爬上樹;而如果是警示有老鷹出現,猴群則會逃往地面。草原犬鼠同樣有類似的機制來提示不同物種、體型與移動速度的掠食者。大象也能夠過使用許多不同的聲音與其他個體進行溝通,包括呼嚕聲、尖嘯、共鳴聲(透過象鼻發出)、吼叫聲與隆隆聲;其中隆隆聲屬於低音頻的次声波,低於人類的聽力範圍,但卻能夠被遠在9.7公里外的其他大象聽見。
哺乳动物的发出信号的方式多种多样。许多动物能发出反捕食者信号,比如鹿与瞪羚可通过四脚弹跳来显示自己的身体状况、逃跑能力,白尾鹿等被捕食者也会在警戒时摇动尾巴,告诫捕食者自己已经觉察到了它的存在。许多哺乳动物会用气味标记自己的领域,有的是出于防御目的,可在种间、种内起到识别作用。小蝙蝠和包括海豚在内的齒鯨能通过声音社交、回声定位。
饮食
哺乳动物要维持恒常体温会消耗很多能量,因此需要大量补充丰富的营养。虽说最早的一批哺乳动物很可能是捕食者,但后来它们逐渐演化,各个不同物种间的饮食结构也有了很大的区别。肉食動物(包括食虫动物)以捕猎、进食其他动物的肉为主。植食动物的饮食以植物为主,会食用纤维素等复合碳水化合物。植食动物还能继续细分下去,如食种动物、食叶动物、食果动物、食蜜动物、食胶动物和食菌动物。食植动物主要在多室胃或盲肠中消化食物。它们的消化道中含有大量细菌,可协助发酵这些复合物质使其易于消化。有些哺乳动物会食粪,撷取糞便中未被吸收的营养物质。雜食動物既吃植物也吃别的动物。肉食动物的消化系统结构较为简单,因为肉类含有的蛋白质、脂類和矿物质不需要特别的消化步骤。不过,鬚鯨的多室胃有腸道菌群,和陆生食植动物类似。
动物体型与其饮食结构也有关系(艾伦法则)。小型哺乳动物的表面积-体型比率大,散热速率也较高,因此需要获取更多的能量来满足其高代谢速率。体重小于510公克的哺乳动物多为食虫动物,因为植物的消化得慢,赶不上它们消耗能量的速度。大型动物能产生更多能量,能量消耗速率也低,因此能够慢慢觅食(使用大型有脊椎动物的肉食动物)或慢慢消化(食植动物)。此外,体重大于510公克的动物通常无法收集到足够多的昆虫支撑其消耗量,大型食虫动物只有那些能一口气吃下整个蚂蚁窝或者白蚁窝尽的能够生存下去。


有的杂食性哺乳动物会更偏向食肉或者食植。鉴于植物和肉类的消化方式不同,哺乳动物在进食时也会更偏爱其中一种,比如有些熊类喜欢吃肉,另一些则更爱吃植物。依照饮食结构的不同,人们将杂食性哺乳动物分为三大类——饮食中肉类占比不到50%的低级食肉动物、占比50%至70%的中级食肉动物和占比70%以上的超级食肉动物。低级食肉动物的牙齿呈钝三角形,方便磨碎食物。超级食肉动物的牙齿由圆锥形牙齿和锋利的裂齿组成,可用于切割食物。有的超级食肉动物(如鬣狗)的下颚十分强壮,可咀嚼骨骼。一些已经灭绝的动物(如剑齿虎亚科)还长有刀刃状的犬齿。
有的哺乳动物虽然生理上是肉食动物,但却会食用植物;还有的反过来,长着植食动物的消化系统但是吃肉。从行为学角度来看这样的动物应该被划入杂食性动物的范畴,不过从生理角度讲,这可能是动物为了保持膳食营养均衡而出现的进食行为。杂食动物必须从肉类、植物中同时吸收微量营养和能量,二者缺一不可。上文提到的会吃肉的植食动物和会吃植物的肉食动物都只是在获取微量营养,它们的能量来源还是遵从它们所在的分类的。许多有蹄类动物,如长颈鹿、骆驼、牛等,会啃咬骨头来获取一些营养素和矿物质。猫固然是食肉动物,但有时也会吃草,帮助它们吐出无法消化的毛球、产生血红蛋白、排便。
无法觅到足够食物时,许多哺乳动物会进入冬眠状态,降低新陈代谢率来减少能量消耗。冬眠前,大型哺乳动物(如熊)会大量进食,在体内屯积大量脂肪,小型哺乳动物则会收集、贮藏大量食物。冬眠期间,新陈代谢速率降低,心跳、呼吸速度放缓,体温甚至能下降到和周遭环境一样的温度。比如,北极地松鼠在冬眠时体温可降至-2.9°C,但头与颈会始终保持在0°C以上。在旱季、酷暑时节,一些生活在燥热环境中的哺乳动物(肥尾鼠狐猴)会夏眠。
智能
在智力較高的哺乳動物中(如靈長類),腦部中大腦的占比也較高。智力本身難以定義,但是有很多行為可以顯示出該個體具有智力,例如學習能力,以及根據情況改變行為的能力。舉例來說,普遍認為大鼠的智力較高,能夠學習並做出各種行為,當大鼠移動到全新的環境中時,這些能力能夠協助適應新環境。某些哺乳類的覓食行為也與智力有關,例如肉食性的貓大腦比植食性的鹿來得大,因為貓必須要智取獵物。

使用工具可以反映出學習能力與認知能力,海獺會用石頭砸開鮑魚的殼,石頭在覓食行為中佔了極重要的部分,研究顯示有些地區的海獺會花上21%的時間製作石頭。黑猩猩會用樹枝來釣白蟻,此行為可能是透過觀察其他個體學習而來,很可能是動物教導行為的案例。有些動物在面對難題時,會經歷尤里卡效應,使用工具來解決難題。其他不會使用工具的動物,例如狗,也會經歷尤里卡效應。
以前學者認為腦容量可以反應出動物智能。大部分的大腦是用於維持身體機能,所以大腦占體重的比例越高,代表用於處理認知能力的部分越多。哺乳動物的大腦是異速生長,其腦化指數大約是 2/3 - 3/4,腦化指數也可用於推估動物智能。抹香鯨的大腦是世界上最大的,成年雄性的大腦體積可達 8,000 立方公分,重達 7.8 公斤。
自我意識是抽象化思考的象徵。雖然自我意識沒有明確的定義,但普遍認為自我意識是後設認知的前提。鏡子測試是傳統上測試動物是否擁有自我意識的方法。某些哺乳類能通過鏡子測試,如亞洲象、黑猩猩、倭黑猩猩、猩猩屬、18個月大的人類、瓶鼻海豚、虎鯨 和偽虎鯨。
社會性
真社會性是最為複雜的社會性,真社會性定義包括世代重疊、階級分工、和共同育幼,大部分的真社會性動都是昆蟲,如蜜蜂、螞蟻,和白蟻,哺乳類之中只有兩種囓齒目為真社會性:裸鼴鼠和達馬拉蘭鼴鼠。


亞社會性動物具有類似於真社會性動物的行為,也就是群居、共同育幼,和原始的階級分工,但其行為無法滿足真社會性的定義。人類和狨亞科(狨屬和檉柳猴屬)的合作育幼行為在靈長類中較為特殊。心理學家哈利·哈洛在1958年設計了一場實驗,以恆河猴(亞社會性靈長類)作為材料,發現了幼猴在心智與性能力發展的過程中,社交是不可或缺的。
分裂融合社群是指大小與組成会不断变化的社群。其中,「母社群」长期稳定存在,包括社群中所有个体。母社群也会分裂出大小、功能各异的“子社群”,这些子社群的规模会随着环境因素、社会事件不断变动。舉例來說,一小群雄性在白天時會脫離母社群(分裂),進行狩獵或尋找食物,夜晚時會回到母社群(融合),分享食物或參與社交活動。很多哺乳動物都具有分裂融合社群,如靈長類、象、斑鬣狗、獅,和海豚。
獨居動物有自己的领地,除了繁殖季以外不會和其他個體進行社會行為。獨居可以避免多个个体在同一地盘上相互竞争食物資源。獨來獨往也比較不易被掠食者或獵物發現。

有些動物具有社会階級制度,個體會爭奪地位。有的动物社会(如狼和狐獴)奉行“專制階級制度”,其中有一個體佔上位,其他個體皆為下位。線型階級制度則是族群內的階級形成線型的关系链,除了最上位者與最下位者以外,每一個體都有其上位者與下位者。有些物種的線型階級制度包括性別,一性別中地位最低的個體,其地位高過於另一性別中地位最高的個體,例如鬣狗。上位者成功繁殖的機率較高,尤其是群體內雌性比雄性多的物種。非群體內的外來個體有時候會被群體接受,進而獲得繁殖的機會,但在某些物種中,此情況極少發生,如吸血蝠。
有些物種具有嚴格的一夫一妻制,一生只有一個伴侶,就算伴侶死去也不會再找新的,例如狼、歐亞河狸,和水獺。除了一夫一妻制以外,還有三種配對組合:(一)一或數雄性上位個體具有繁殖權的一夫多妻制;(二)雌性個體和數名雄性個體交配的一妻多夫制;(三)數個雄性個體與數個雌性個體交配的多夫多妻制。不考量求偶場的情況下,90% 的哺乳類都是一夫多妻制。求偶場是指數名雄性圍繞在雌性旁邊並對其求偶,如港海豹。
胎生和哺乳是所有的高等哺乳類(单孔目除外)都有的繁殖特徵。育幼行為則因物種而異,有些物種築巢,有些物種掘洞,有些物種則會長時間照顧後代。大部分哺乳類是K策略,也就是在後代上投資更多的時間和能量,取而代之的是單次繁殖能產生後代數量較少,而R策略則相反。雄性與雌性都可以從繁殖行為中獲利,然而在大多數的情況下,兩方的獲利是不對等的。有些物種的雌性具有護幼行為,會攻擊其他雌性與幼獸,然而,有些物種會協助親戚育幼。有些物種的雄性也會育幼,如马岛猬属,然而這個行為因物種而異,就連同個屬內的物種育幼行為都有可能不同,舉例來說,日本獼猴的雄性會育幼,但是豚尾獼猴的雄性不會。
人类与其他哺乳动物
人类文化中的哺乳动物

人类以外的哺乳动物在文化中扮演了各式各样的角色。哺乳动物是世界上最受欢迎的宠物——全世界饲养的宠物猫、狗、兔、鼠数量成千上万。其他哺乳动物,如猛犸象、马、鹿等,则是人类最古老的艺术主题,拉斯科洞窟壁画等旧石器时代晚期的岩画中会见到它们的身影。许多画家都以画哺乳动物出名,像阿尔布雷希特·丢勒(画过犀牛)、乔治·斯塔布斯(擅画马)、埃德英·藍道西爾(擅画红鹿)等等。人类会出于获取食物或体育运动的目的狩猎其他哺乳动物,其中鹿和野猪是最常见的猎物。像赛马、赛狗这样的哺乳动物则用于体育竞技,通常与赌博活动有关。关于能否将动物作为人类的伴侣带在身旁,抑或应当保障动物的权利、让动物独自生存,人们至今仍未达成共识。哺乳动物也在文学、电影、神话、信仰中广泛现身。
对哺乳动物的利用

世界上的肉用家畜中,驯化的哺乳动物占了绝大多数。其中包括(2009年)约14亿头牛、10亿隻绵羊、10亿头家猪、和(1985年)逾7亿只兔子。包括牛和马匹在内的役用動物都被用作农业工作和运输,但随着机械化运输和农业机械的到来,它们的数量有所下降。2004年,这些动物为第三世界的小型农场提供了约80%的功率输出,涉及到世界范围内约20%的运输吞吐量(主要集中在农村)。在车辆寸步难行的山区,驮畜仍是货运的主力军。哺乳动物的皮還被制成皮革,成为鞋、服装和室内装潢的原材料。羊毛也来自哺乳动物,其中包括绵羊、山羊和羊驼,几个世纪以来一直用来做衣服。
哺乳动物作为实验动物在科学中发挥着重要作用,常常用在遗传学的基础生物学研究和在新药的开发中。新药必须经过彻底测试,以证明其安全性。每年有百万哺乳动物,特别是老鼠,被人类用于实验。科学家会将小鼠的基因人工修改,植入人工DNA片段或破坏一段DNA,这样可让部分基因失活。这样处理过的实验小鼠被称为“基因剔除小鼠”,能协助科学家研究功能未知的基因,从而对基因进行测序。还有一小部分实验哺乳动物是非人类灵长类动物,主要被用来研究与人类的相似度。
查尔斯·达尔文和賈德·戴蒙等人已经注意到哺乳动物在新石器时代的农业和文明发展中的重要性。人工驯养的哺乳动物导致世界各地的农民大范围取代了狩猎采集者。从狩猎采集到遊牧,再到农业社会,哺乳动物帮助人类跨越了一个又一个社会发展的里程碑。以驯养哺乳动物为基础的新农业经济造成了“人类社会的全面重组、全球生物多样性的变化以及地貌及大气层的重大改变……等影响深远的事件”。
杂交


两个遗传上有差异的个体杂交产生的后代即杂交种,杂交种时常(但不绝对)具有高度杂合性。在人类圈养的动物中有意或无意的杂交活动已持续了几千年,并带来经济利益。物种亚种之间的杂交称为种内杂交,如孟加拉虎与东北虎。同属不同物种间的杂交称为种间杂交,如狮子和老虎。不同属物种间的杂交称为属间杂交,如绵羊和山羊。在杂交带,同属或同种动物生活在相同或毗邻区域,会发生自然杂交。一些杂交种被视为物种,如紅狼(有争议)。
人工选择繁殖家养动物正被用于“恢复”近期灭绝的动物,其目标是获得表型类似于灭绝野生型祖先的种系。外观是逆向育种的重要目标,德国的原牛逆向育种实验导致了外观上较为相似的赫克牛。使用种内杂交进行逆向育种获得的种系可能在外观、生态位以及部分基因上与野生型十分相似,但野生型的初始基因庫已随着灭绝永久消失了。
来自家养动物的基因污染、不受控的杂交、基因滲入会使野生种群与家养种群同质化,这将导致生活在特定生态环境中的純種野生种群受到灭绝威胁。一些种群,特别是稀有种系的灭绝,可能发生于新种群的引入、选择性繁殖动物、或栖息地变化导致分离的种群相互接触之时。虽说自然演化也伴随着一定程度的基因流動,受人类影响的雜交会污染稀有纯种种群的基因库,而且基因污染直至灭绝的现象在生物形態學上不总容易观察。濒危的河水牛已因家养水牛的基因污染受进一步灭绝威胁。
威胁

人类活动是动物灭绝的主要原因,导致部分生态环境缺少大型脊椎动物(即空森林现象)。在第四紀滅絕事件中,巨型動物群的大量滅絕恰好約與現代人類的出現時間重疊。有假说认为,人類曾大批獵殺真猛獁象等大型哺乳動物,使牠們的數量锐减進而滅絕。2019年,IPBES在《全球生物多樣性與生態系服務評估報告》中提到野生哺乳動物的總生物量自人類文明出現至現在已減少了82%。
許多物種被認為於不遠的未來即會滅絕,包括犀牛、數個灵长目物種、穿山甲、长颈鹿等。光是人類的狩獵行為就導致了全球數百種哺乳動物面臨滅絕危機,肉类食物的需求增加被認為是促成森林砍伐與棲息地破壞的背後主要原因,並因此導致生物多樣性減少,在一些物種豐富的棲息地,例如亞馬遜雨林,許多地區已被人類破壞並逐漸開發轉為生產肉類的農牧用地。根據2016年世界自然基金会的地球生命力指数,自1970年以來全球野生動物的數量已經減少了58%,主因包括棲息地破壞、過度捕獵及汙染。如果按照目前的趨勢繼續下去,於2020年時將會有67%的野生動物消失。另一個對物種生存造成威脅的原因是過度捕獵與盜獵,導致被作為獵物的動物數量減少,尤其是那些棲息於村莊附近的物種,例如西猯。其他捕獵的原因包括獲取牠們身上的資源,例如象牙交易,就直接導致了非洲象數量的減少。除此之外,誤捕與廢棄漁具纏繞也對海洋哺乳動物的生存造成威脅,以鯨豚為例,每年約有65,000至86,000頭個體因此而死亡。
目前的瀕臨滅絕物種正受到全球的關注,主要是透過《生物多樣性公約》(又稱為《里約公約》),由196個國家共同簽屬,致力於推動並落實公約的三大目標,包括保育生物多樣性、永續利用其組成分子,以及公平合理地分享生物多樣性遺傳資源所產生的利益。另一個著名的保育組織為國際自然保護聯盟,其中的會員包括了超過1,400個政府與非政府组织。
近代哺乳動物的滅絕幾乎都是受到人類的影響而滅絕,其中國際自然保護聯盟界定「近代滅絕」 是指西元1500年以後才滅絕的動物,從該時算起至2015年,已有約80種哺乳動物滅絕。有些物種,如麋鹿,為野外绝灭,目前僅存有人工飼養的個體。其他物種,如佛羅里達山獅,為生態滅絕,牠們的數量過於稀少,以至於已經無法對當地生態系造成任何影響。其他物種則有不少為局部地區滅絕,雖然還未滅絕,但分布地區已大幅萎縮,例如現今灰鯨就已經於大西洋局部滅絕。
注解
参见
参考资料
拓展阅读
- Brown WM. Natural selection of mammalian brain components. Trends in Ecology and Evolution. 2001, 16 (9): 471–473. doi:10.1016/S0169-5347(01)02246-7.
- Jaffa Kv, Taher NA. Mammalia Palaestina: The Mammals of Palestine. The Palestinian Biological Bulletin. 2006, (55): 1–46.
- McKenna MC, Bell SK. Classification of Mammals Above the Species Level. New York: Columbia University Press. 1997. ISBN 978-0-231-11013-6. OCLC 37345734.
- Nowak RM. Walker's mammals of the world 6. Baltimore: Johns Hopkins University Press. 1999 [2021-06-25]. ISBN 978-0-8018-5789-8. OCLC 937619124. (原始内容存档于2022-06-01).
- Simpson GG. The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History. 1945, 85: 1–350.
- Murphy WJ, Eizirik E, O'Brien SJ, Madsen O, Scally M, Douady CJ, et al. Resolution of the early placental mammal radiation using Bayesian phylogenetics. Science. 2001-12, 294 (5550): 2348–51. Bibcode:2001Sci...294.2348M. PMID 11743200. doi:10.1126/science.1067179.
- Springer MS, Stanhope MJ, Madsen O, de Jong WW. Molecules consolidate the placental mammal tree (PDF). Trends in Ecology & Evolution. 2004-08, 19 (8): 430–8 [2020-07-04]. PMID 16701301. doi:10.1016/j.tree.2004.05.006. (原始内容存档 (PDF)于2016-07-29).
- Vaughan TA, Ryan JM, Capzaplewski NJ. Mammalogy 4. Fort Worth, Texas: Saunders College Publishing. 2000. ISBN 978-0-03-025034-7. OCLC 42285340.
-
Kriegs JO, Churakov G, Kiefmann M, Jordan U, Brosius J, Schmitz J. Retroposed elements as archives for the evolutionary history of placental mammals. PLoS Biology. 2006-04, 4 (4): e91. PMC 1395351
 . PMID 16515367. doi:10.1371/journal.pbio.0040091.
. PMID 16515367. doi:10.1371/journal.pbio.0040091.
- MacDonald DW, Norris S. The Encyclopedia of Mammals 3. London: Brown Reference Group. 2006. ISBN 978-0-681-45659-4. OCLC 74900519.
外部链接
| 維基教科書中的相關電子教程:哺乳动物 |
- Biodiversitymapping.org – 世界上所有哺乳动物的分布图 (页面存档备份,存于互联网档案馆)
- Paleocene Mammals (页面存档备份,存于互联网档案馆), 一个关于哺乳动物诞生的网站, paleocene-mammals.de
- Evolution of Mammals (页面存档备份,存于互联网档案馆), 早期哺乳动物的简介, enchantedlearning.com
- European Mammal Atlas EMMA (页面存档备份,存于互联网档案馆) 欧洲哺乳动物协会, European-mammals.org
- Marine Mammals of the World (页面存档备份,存于互联网档案馆) – 所有海洋哺乳动物的概述,包括全水生哺乳动物和半水生哺乳动物, noaa.gov
- Mammalogy.org (页面存档备份,存于互联网档案馆) 美国哺乳动物学家协会,成立于1919年,旨在促进哺乳动物的研究,网站包括一个哺乳动物图像库
|
|