
| HIV-1蛋白酶(逆转录病毒氨酰蛋白酶) | |||||||
|---|---|---|---|---|---|---|---|

| |||||||
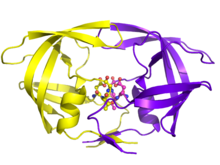
| HIV-1 蛋白酶二聚体为白色和灰色,肽底物为黑色,活性位点天冬氨酸侧链为红色。 (PDB 1KJF) | |||||||
| |||||||
| 识别码 | |||||||
| EC編號 | 3.4.23.16 | ||||||
| CAS号 | 144114-21-6 | ||||||
| 数据库 | |||||||
| IntEnz | IntEnz浏览 | ||||||
| BRENDA | BRENDA入口 | ||||||
| ExPASy | NiceZyme浏览 | ||||||
| KEGG | KEGG入口 | ||||||
| MetaCyc | 代谢路径 | ||||||
| PRIAM | 概述 | ||||||
| PDB | RCSB PDB PDBj PDBe PDBsum | ||||||
| 基因本体 | AmiGO / EGO | ||||||
| |||||||
HIV-1蛋白酶是一种逆转录病毒天冬氨酰蛋白酶(retropepsin),它是一种参与逆转录病毒肽键水解的酶,它对导致艾滋病的逆转录病毒,HIV的生命周期至关重要。HIV蛋白酶在九个切割位点切割新合成的多蛋白(即Gag和Gag-Pol)以产生HIV病毒粒子的成熟蛋白质成分,这是病毒在宿主细胞外的感染形式。如果没有有效的HIV蛋白酶,HIV病毒粒子将保持无感染性。
结构
成熟的HIV蛋白酶以22 kDa同型二聚体形式存在,每个亚基由 99 个氨基酸组成。单个活性位点位于相同的亚基之间,并具有天冬氨酸蛋白酶共有的特征性Asp-Thr-Gly(Asp25、Thr26 和 Gly27)催化三联体序列。由于HIV-1蛋白酶只能作为二聚体发挥作用,因此成熟的蛋白酶包含两个Asp25氨基酸,一个来自每个单体,它们作为催化残基相互结合。此外,HIV蛋白酶有两个分子“襟翼”,当酶与底物结合时,它们移动的距离可达7Å。这可以通过襟翼打开和关闭的动画来可视化。
生物合成

前体
Gag-Pol多聚蛋白,其中包含过早的编码蛋白,也包括HIV-1蛋白酶。HIV-1蛋白酶位于反转录酶(位于 PR 的 C 端)和转框区(Transframe region,TFR)的p6pol(位于 PR 的 N 端)之间。
为了使该前体成为功能性蛋白质,每个单体必须与另一个HIV-1蛋白酶单体结合,通过各自贡献各自催化三联体的Asp25来形成功能性催化活性位点。
合成机制
当病毒性HIV-RNA进入细胞时,他将会被逆转录酶、整合酶和成熟的HIV-1蛋白酶伴随。逆转录酶将病毒性RNA转化为DNA,促进整合酶在将病毒遗传信息与宿主细胞DNA结合中的作用。病毒性DNA可以在细胞核中保持休眠状态或者也可以转录成mRNA并由宿主细胞翻译成Gag-Pol多聚蛋白,然后成熟的HIV-1蛋白酶将会将其切割成单个功能蛋白(包括新合成的HIV-1蛋白酶)。
HIV-1蛋白酶前体通过促进其从Gag-Pol多聚蛋白中的切割以一种称为自动加工的机制来催化其自身的产生。HIV-1蛋白酶的自动加工以两个连续步骤为特征:(1)在p6pol-蛋白酶切割位点对N端进行分子内切割,这有助于完成HIV-1蛋白酶加工并增加新形成的蛋白酶-逆转录酶中间体的酶活性。(2)C末端在蛋白酶-逆转录酶切割位点的分子间切割,导致两个HIV-1蛋白酶亚基组装成成熟的二聚体。两个亚基的二聚化允许形成功能齐全的组合活性位点,其特征在于两个Asp25催化残基(每个单体一个)。


功能
HIV-1蛋白酶具有两个目的。前体HIV-1蛋白酶负责通过HIV-1蛋白酶自动加工催化其自身产生成熟的HIV-1蛋白酶。成熟的蛋白酶能够水解Gag-Pol多聚蛋白上九个特定位点的肽键,将产生的亚基加工成成熟的、功能齐全的蛋白质。这些切割的蛋白质,包括逆转录酶、整合酶和RNaseH,由病毒复制所必需的编码区成分编码。
机制
作为一种天冬氨酸蛋白酶,二聚化的HIV-1蛋白酶通过天冬氨酰基团复合物进行水解。在HIV-1蛋白酶的联合催化活性位点上的两个Asp25残基中,一个被去质子化而另一个被质子化,这是由于与微环境的pKa差异。
在一般的天冬氨酸蛋白酶机制中,一旦底物与酶的活性位点正确结合,去质子化的Asp25催化氨基酸就会经历碱催化,从而通过去质子化来使进入的水分子成为更好的亲核试剂。产生的氢氧根离子攻击肽键的羰基碳,形成一个具有瞬态氧阴离子的中间体,该中间体由最初质子化的Asp25来稳定。氧阴离子重新形成双键,导致两个氨基酸之间的肽键断裂,而最初去质子化的Asp25经过酸催化将其质子提供给氨基,使氨基成为更好的离去基团,以完成肽键断裂并恢复到原来的去质子化状态。
虽然HIV-1蛋白酶与非病毒天冬氨酸蛋白酶具有许多相同的特征,但一些证据表明HIV-1蛋白酶以协同方式催化水解。换句话说,亲核水分子和质子化的Asp25在催化过程中同时攻击易断裂的肽键。

作为药物靶点
凭借其在HIV复制中不可或缺的作用,HIV蛋白酶已成为主要的药物治疗。 HIV蛋白酶抑制剂通过模仿其底物的四面体中间体特别是来结合活性位点并基本上“卡住”,使酶失效。在组装和出芽后,缺乏活性蛋白酶的病毒颗粒不能成熟为感染性病毒粒子。几种蛋白酶抑制剂已获准用于HIV治疗。
有10种目前已获美国食品和药物管理局批准的HIV-1蛋白酶抑制剂:茚地那韦、沙奎那韦、利托那韦、奈非那韦、洛匹那韦、安普那韦、福沙那韦、阿扎那韦、替拉那韦和达芦那韦。许多抑制剂具有不同的分子成分,因此具有不同的机制作用,例如阻断活性位点。它们的功能作用还延伸到影响其他抑制剂药物(利托那韦)的循环浓度,并且仅用于病毒对其他抑制剂(替拉那韦)表现出耐受性的某些情况。
进化与耐药性
由于逆转录病毒的高突变率,特别是由于突变敏感区域(尤其是包含催化三联体序列的区域),并且考虑到 HIV蛋白酶中一些氨基酸的变化可以使其对抑制剂的明显性大大降低,因此在复制抑制药物的选择压力下,这种酶的位点会迅速改变。
两种类型的突变通常与增加的耐药性有关:“主要”突变和“次要”突变。主要突变涉及HIV-1蛋白酶活性位点的突变,阻止选择性抑制剂与其结合。次要突变是指由于长期暴露于类似化学物质从而导致酶外围的分子变化,可能影响HIV-1蛋白酶的抑制剂特异性。
将HIV耐药性的发展降至最低的一种方法是同时使用一种药物组合,这些药物可以同时抑制HIV复制周期的几个关键方面,而不是一次使用一种药物。其他药物治疗靶点包括逆转录酶、病毒附着、膜融合、cDNA整合和病毒粒子组装。
| |||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||