
| 白色念珠菌 | |
|---|---|
| |
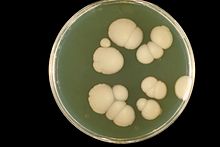
| 生長於沙鮑弱氏瓊脂的白色念珠菌 | |
|
科学分类 | |
| 界: | 真菌界 Fungi |
| 门: | 子囊菌门 Ascomycota |
| 纲: | 酵母菌綱 Saccharomycetes |
| 目: | 酵母菌目 Saccharomycetales |
| 科: | 酵母菌科 Saccharomycetaceae |
| 属: | 念珠菌屬 Candida |
| 种: | 白色念珠菌 C. albicans |
| 二名法 | |
|
Candida albicans | |
| 異名 | |
| |
白色念珠菌(學名:Candida albicans)是一種能造成伺機性感染的酵母菌,常見於人類消化道與泌尿生殖道的菌群,約有四成至六成健康成人的口腔與消化道中都有白色念珠菌,平時與人體行片利共生,但可在人體免疫缺陷時過度生長而造成念珠菌症,是念珠菌屬最常見的致病菌種,與熱帶念珠菌、近平滑念珠菌、光滑念珠菌和克魯斯念珠菌合計造成了95%的念珠菌症感染,其中白色念珠菌便占了66%,但此比例正持續下降中。白色念珠菌造成的感染有口腔、食道與陰道等處的局部粘膜感染,也有嚴重的全身性感染,有研究指出後者的致死率高達40%,另有一項統計顯示白色念珠菌造成的院內感染在美国每年造成2,800至11,200人死亡。白色念珠菌也是最常在醫療器材上形成生物薄膜的真菌,生物薄膜中的白色念珠菌抗性比游離成長者還高,可隨著醫療器材入侵血液而造成嚴重感染。
白色念珠菌多以雙倍體的型式存在,不同於多數真菌為單倍體,但近年漸有研究發現少數情況下白色念珠菌也可形成單倍體與四倍體,其雙倍體的基因組長度為29Mb,另外其基因組中,CUG密碼子對應的胺基酸為絲胺酸,與絕大多數生物中對應的白胺酸不同,是相當罕見的遺傳密碼例外情形。白色念珠菌常被當作真菌感染研究的模式生物,容易在實驗室中培養,並可以改變培養基成分的方式控制其生長型態。白色念珠菌屬於雙態性真菌,可以酵母型或菌絲型兩種方式生長,除此之外還可在多種不同的形態表型間互相轉換,如白色型-混濁型轉換可以改變菌落的外形與基因表現,進而促進準性生殖。
名稱與分類
白色念珠菌的學名Candida albicans為一同義反覆,其中屬名Candida來自拉丁文的candidus,意指白色,種小名albicans來自拉丁文的albicō,意思亦為白色。早在西元前400年,希波克拉底在其著作《論流行病》(Of the Epidemics)中即提到了鵝口瘡,可能是白色念珠菌感染的最早記載。
白色念珠菌的同物異名有多達上百個,最早於1853年由法國生物學家查爾斯-菲力毗·羅賓發表描述,命名為Oidium albicans,1890年,德國真菌學家弗里德里希·威廉·措普夫調整其分類,改其學名為Monilia albicans(其中Monilia來自拉丁文的monile,意指項鍊)。1923年,荷蘭真菌學家克里斯汀·瑪麗·別爾克豪特發表了新屬念珠菌屬,並將白色念珠菌歸入該屬,改為現名Candida albicans。
種系發生
已有許多分子種系發生學的研究以不同數量、種類的基因序列分析念珠菌屬中各物種的親緣關係。念珠菌屬中,與白色念珠菌關係最近的物種為都柏林念珠菌(C. dubliniensis),兩者組成的演化支與熱帶念珠菌(C. tropicalis)互為姊妹群,而三者組成的演化支又與近平滑念珠菌(C. parapsilosis)和长孢洛德酵母(Lodderomyces elongisporus)組成的演化支互為姊妹群,以上類群均以雙倍體的形式存在,不同於多數真菌為單倍體,顯示其共祖可能發生了單倍體-雙倍體的轉換。這些雙倍體的類群與其他單倍體的類群(季也蒙念珠菌、葡萄牙念珠菌與汉逊德巴利酵母)共同組成念珠菌演化支(Candida clade),以上類群均將密碼子CUG編碼為絲胺酸,而不像絕大多數生物將其編碼為白胺酸,顯示念珠菌演化支的共祖可能發生了一次CUG密碼子對應的tRNA重组。念珠菌屬並非單系群,同屬念珠菌屬的光滑念珠菌不屬於念珠菌演化支,而是與釀酒酵母等酵母菌關係較近,共同組成酵母菌演化支(Saccharomyces clade),與其他念珠菌的親緣關係較遠。
|
|||||||||||||||||||||||||||||||||||||||||||||||||
基因组
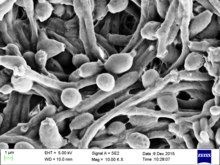
单倍体的白色念珠菌的基因组的大小約为15Mb(雙倍體約為29Mb),由8條染色体对组成,二倍體中其中一組染色體分別称为chr1A、chr2A、chr3A、chr4A、chr5A、chr6A、chr7A和chrRA,另一組染色體具有相似的名称,但最后一个字為B,如chr1B、chr2B、...和chrRB。整个基因组包含6198个开放阅读框(ORF),截至2019年2月,这些ORF中仍有70%尚未被鉴定。有研究使用四環素轉錄調控系統控制白色念珠菌各個基因的表現,建構了一個GRACE(基因置换和条件表达)資料庫,研究白色念珠菌基因组中各基因對其生長的必要性,結果顯示其中有567個基因是其生長所必須的。研究白色念珠菌最常用的菌株是WO-1和SC5314菌株,WO-1菌株可高频率地在白色型(white)與混濁型(opaque)之间互相轉換,而SC5314菌株是用作基因組参考序列的菌株。白色念珠菌两种常用菌株的基因组已經完成全基因組定序,其中SC5314菌株完成于2004年,是较早完成测序的真菌之一。
白色念珠菌核基因組的一大特色是密碼子CUG對應的胺基酸不像其他生物中為白胺酸,而是絲胺酸,同屬念珠菌演化支的其他物種亦有此現象,這種遺傳密碼發生例外的情形在真核生物的细胞核基因组中是相當罕見的,在原核生物中也仅出現在起始密碼子。此現象可能使白色念珠菌長期處在熱休克反應的狀態,有助於提升其對逆境的抗性。有研究分析比較白色念珠菌與釀酒酵母基因中的CUG密碼子,發現白色念珠菌的CUG密碼子(編碼絲胺酸)在釀酒酵母的同源序列中大多是對應編碼絲胺酸與其他親水胺基酸的的密碼子,而釀酒酵母的CUG密碼子(編碼白胺酸)在白色念珠菌的同源序列中大多是對應編碼白胺酸與其他疏水胺基酸的密碼子,顯示CUG密碼子功能的改變并未对蛋白质的构象造成太大变化。有研究将白色念珠菌识别CUG密碼子的tRNA导入到釀酒酵母中表現,使其轉譯時隨機將部分CUG密碼子辨認為對應絲胺酸,結果啟動了抗逆境反應,造成酵母維持在二倍體的狀態,且抑制了交配。非標準的遺傳密碼使白色念珠菌蛋白質交互作用的研究難以在模式生物釀酒酵母中進行,對此已有研究團隊開發了白色念珠菌的雙雜合系統。
白色念珠菌的基因组有高度杂合性,有着大量的SNP位点,常因染色体長度多型性(重複序列的長度不一)、相互易位、染色體刪除與個別染色體成為三體等因素,造成染色體數量或結構上的變異,諸如此類核型的改变會导致表型的改变,是這種真菌對不同環境适应的機制之一。随着白色念珠菌基因组資訊的成功解碼,關於這些機制的研究正持續進行中。
研究工具
白色念珠菌是用來研究真菌感染的重要模式生物,許多研究團隊開發了白色念珠菌的各種資料庫與其分子生物學研究的工具。2004年,多位研究者合作開發了念珠菌基因組資料庫(Candida Genome Database, CGD),該資料庫是由酵母菌基因組資料庫修改而來,許多開發者過去也曾參與酵母菌基因組資料庫的構建。念珠菌基因組資料庫最初提供白色念珠菌SC5314菌株的基因組資訊,以全基因組霰彈槍定序法定序。後來也陸續加入白色念珠菌的其他菌株與念珠菌屬的其他物種。
對白色念珠菌進行基因轉殖後,常用的選擇性標記包括CaNAT1(抗諾爾絲菌素)、MPAr 與IMH3r(抗黴酚酸)等,也有使用營養缺陷型者,例如URA3基因的產物為乳清酸核苷-5'-磷酸脱羧酶,是合成單磷酸尿苷所需的酵素,為白色念珠菌生長所必須,因此URA3營養缺陷型的突變株只有在成功轉殖帶有URA3的質粒後才能在不含尿嘧啶的特殊培養基中生長,URA3因而可作為基因轉殖後的選擇性標記,但許多研究顯示轉殖的URA3從質粒整合進染色體上的位點不同,會使其表現量不同而影響白色念珠菌的致病能力,而可能造成研究中判斷轉殖基因功能的困難,為此已有研究團隊開發組胺酸(HIS1)、白胺酸(LEU2)與精胺酸(ARG4)營養缺陷型的菌株與相應質粒,這些基因的轉殖不會影響白色念珠菌的致病能力,以避免使用基因轉殖研究特定基因對致病能力影響時,對研究結果造成干擾。
以質粒進行基因轉殖時,常用於釀酒酵母的質粒在白色念珠菌中多有在染色體外不穩定或基因表達效率低下的問題,有研究團隊開發了適用於白色念珠菌基因轉殖的CIp10(Candida integrating plasmid 10)質粒,以核糖體蛋白RP10基因為整合進染色體中的同源序列,並帶有用作選擇性標記的URA3基因,因其整合進染色體的位置固定,可避免URA3表現量因整合位點不同而有異,進而影響致病能力。CIp10質粒可以高效率的轉殖進白色念珠菌中,並有多個研究團隊對其進行微調,設計出各種衍生的質粒。
型態

白色念珠菌有許多不同的形態表型,除了作為雙態性真菌,可以在酵母菌型態與菌絲型態間轉換之外,還可以進行通稱為「高頻率轉換」(high-frequency switching)之多種其他表型間的轉換,其中被研究最多的一種轉換則是某些菌株白色型-混濁型間的轉換,另外還有許多其他不同的轉換系統,例如SC5314菌株可在七種表型間互相轉換。酵母菌型-菌絲型轉換的過程相當快速,且通常整個菌落的所有細胞都會一起轉換;高頻率轉換則可能只在菌落中的部分細胞中發生,且經轉換後的細胞通常仍能進行酵母菌型-菌絲型轉換。這些轉換均為可逆,且常受到二氧化碳濃度、氧氣濃度、培養基種類與溫度等環境因子的影響。
酵母菌型─菌絲型轉換
白色念珠菌為雙態性真菌,可以酵母菌或菌絲的型式生長,其中酵母菌型生長時可進行出芽生殖,有時出芽產生的子細胞沒有立即脫離,而形成類似菌絲的絲狀外觀,稱為假菌絲(pseudohypha)。白色念珠菌在實驗室常用的標準培養液生長時通常呈酵母菌型,呈卵圓形,長約6微米,不過溫度、二氧化碳濃度、養分與酸鹼值的些許改變便可能使其轉換成菌絲型,在模擬人體生理環境的培養基中生長時亦呈菌絲型。在適合其生長的環境下,白色念珠菌菌絲萌發時直徑約為2.6微米,成熟菌絲直徑則約為3.4微米。兩種型態在白色念珠菌感染人體時各有功能,酵母菌型較適合在血液中散播,菌絲型則有助於穿透組織、在器官中著生、躲避巨噬細胞的攻擊、以及在醫療器材上形成生物薄膜,這種型態轉換與其致病力密切相關,不過仍有研究指出有無法進行此轉換的菌株亦可進行感染。以小分子藥物抑制白色念珠菌的酵母菌型─菌絲型轉換可能有助於抑制其感染。
另外白色念珠菌還在環境條件惡劣時,可以形成對逆境抗性較高的厚垣孢子。厚垣孢子多著生於從菌絲分枝出去之瓶狀突起的端點。



高頻率轉換
除了酵母菌型-菌絲型轉換外,白色念珠菌亦可進行高頻率轉換(high-frequency switching),在多種表型間相互轉換,這種轉換常是自然發生,並非由特定環境因子調控,且只發生在菌落中的部分細胞。高頻率轉換最早於1985年由美國生物學家大衛·索爾的團隊在SC5314菌株中發現,這種菌株可以在平滑(smooth)、星狀(star)、不規則皺紋狀(irregular wrinkle)、環狀(ring)、點狀(stipple)、帽狀(hat)與絨毛狀(fuzzy)等七種表型間進行轉換,不同表型間轉換發生的頻率各異,介於萬分之一至十分之一之間,且紫外線的照射可大幅提升轉換發生的頻率。高頻率轉換可能與某些基因表現的改變有關,有研究發現缺乏組蛋白脫乙醯酶SIR2的菌株出現高頻率轉換的機率大幅上升,顯示控制轉換的基因平時可能為SIR2所抑制。在各種表型間轉換的能力有助於白色念珠菌在不同環境下的生長。
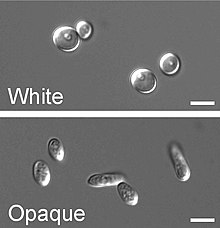
白色型─混濁型轉換
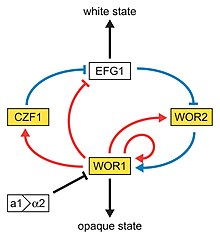
白色型─混濁型轉換(white-to-opaque switching)是白色念珠菌第二種被發現的高頻率轉換,於1986年由大衛·索爾團隊在WO-1菌株中發現,其中白色型(white)的細胞呈圓形,形成白色、平滑的菌落,混濁型(opaque)的細胞形狀則較長,形成灰色且較為扁平的菌落。此轉換是由WOR1(White to Opaque Regulator 1)蛋白調控,WOR1蛋白則受MTL基因調控,當MTLa與MTLα基因同時存在時,會產生a1-α2複合體,抑制WOR1蛋白的表現,進而抑制白色型─混濁型的轉換,又因混濁型是發生準性生殖的主要表型,進行準性生殖的機率比白色型高上百萬倍,自然環境中有高達95%的白色念珠菌都屬MTLa/α型,因此難有此轉換與交配發生。
白色型─混濁型轉換受到許多環境因子調控,其中氧化壓力會促進白色念珠菌轉為混濁型,溫度的提升則會促進其轉為白色型,因此在宿主體內(攝氏37度)生長時白色念珠菌多為白色型,但缺氧、二氧化碳與N-乙酰葡糖胺濃度高的環境(例如消化道中)可以亦可刺激白色型─混濁型轉換,使白色念珠菌呈混濁型,因此感染宿主的白色念珠菌亦可能發生交配。另外當白色念珠菌生長速率降低時,可能因細胞中得以累積足夠濃度的WOR1蛋白,不因細胞分裂而快速被稀釋,進而促進白色型─混濁型轉換。白色型─混濁型轉換對白色念珠菌的致病力也有關聯。白色型較適於體內的全身性感染,混濁型則較適於皮膚的淺表感染,兩種型態對宿主免疫系統的反應與耐受性也有所不同,可能有助於感染。
除了白色念珠菌外,同為念珠菌屬且亦可感染人體的都柏林念珠菌與熱帶念珠菌也能進行白色型─混濁型轉換。2014年,有研究發現除了白色型與混濁型外,還有一種「灰色型」可與兩型態相互轉換,灰色型的白色念珠菌造成皮膚感染的能力比另外兩種型態還強,發生交配的機率則介於兩者之間,且白色型─灰色型的轉換不需WOR1的參與。三種型態各有不同的基因表現,造成其型態、代謝、致病能力、適應宿主體內的環境與對逆境的抗性皆有所不同。
其他轉換
除了白色型─混濁型轉換外,還有許多新的白色念珠菌表型轉換機制陸續被發現,分別存在不同菌株中。從人體陰道中分離的部分菌株可以在菌絲量多與菌絲量少的菌落型態之間互相轉換。SC5314菌株還可進行白色型─消化道型的轉換,消化道型(Gastrointestinally-IndUced Transition, GUT)是存在宿主消化道中的類型,其外型與混濁型相似,細胞形狀較長、形成的菌落較扁,且從白色型轉換的過程亦需WOR1蛋白參與,但以電子顯微鏡觀察時可見其表面沒有混濁型常有的突起,且基因表現亦與混濁型不同,並因維持MTLa/α的基因型而不像混濁型可以進行交配。消化道中可能含有刺激白色型白色念珠菌轉換為消化道型的訊息,此型的白色念珠菌主要在宿主消化道中行片利共生,不對宿主健康造成負面影響。
生殖

白色念珠菌過去被認為只能以有絲分裂進行無性生殖,不能進行減數分裂和產生孢子,沒有以有性生殖繁殖的途徑。現代研究則發現白色念珠菌亦可進行準性生殖,但僅限於MTLa或MTLα基因型的個體,MTLa/α者因會抑制白色型-混濁型轉換而難以交配(詳見#白色型─混濁型轉換一節)。混濁型的MTLa與MTLα的個體可分別釋放MFa與MFα費洛蒙,與彼此細胞表面的受體結合而啟動停止細胞週期、改變細胞型態等交配機制,並可形成一種稱為性菌膜(sexual biofilm)的生物薄膜,有助於交配進行。最終兩細胞可進行核融合而形成四倍體。四倍體在某些環境下不穩定,有絲分裂時可能發生染色體不分離而產生非整倍體,進而在多次有絲分裂後回復為二倍體,其中染色體缺失的過程有協同效應,即丟失數個染色體後,在後續的有絲分裂中繼續丟失其他染色體的機率會提升。有絲分裂時Spo11蛋白可以切割DNA,促使同源染色體間發生同源重組。
白色念珠菌因為雙倍體,其準性生殖為二倍體-四倍體-二倍體的過程,而不同於其他真菌的單倍體-二倍體-單倍體。不過白色念珠菌的二倍體亦可能丟失染色體而形成單倍體,單倍體的a與α菌株亦可交配而形成二倍體。
致病機制
白色念珠菌可以分泌天冬胺酸蛋白酶(secreted aspartic proteinases, Saps)、磷脂酶與脂酶,可分解宿主細胞的胞外基質與細胞膜表面物質,有助其黏著、侵入宿主的組織中,甚至可能可以用以破壞免疫細胞。2016年,有研究發現白色念珠菌感染宿主粘膜時,從酵母菌型轉為菌絲型後,會分泌一種稱為念珠菌素的毒素,念珠菌素長31個胺基酸,是由Kex2p蛋白酶切割Ece1p產生的一種多肽,可造成宿主上皮細胞的損傷,並刺激宿主的免疫反應。
酵母菌型-菌絲型轉換對白色念珠菌的致病能力相當重要。吞噬細胞是宿主抗真菌免疫反應中的重要環節,白色念珠菌被巨噬細胞內吞後,並由酵母菌型轉換為菌絲型,最終穿透巨噬細胞而重新回到組織中。在巨噬細胞內部,白色念珠菌會啟動許多基因的表現,改變其代謝與促進型態轉換,其中許多基因為念珠菌所特有,釀酒酵母中不存在同源基因者,很可能與白色念珠菌的致病能力有關。另外菌絲型的白色念珠菌菌絲表面有Hwp1(Hyphal wall protein 1)蛋白,該蛋白可與宿主上皮細胞表面的轉麩醯胺酶結合,有助白色念珠菌黏附於宿主細胞,進而造成感染。
生物薄膜

白色念珠菌還可形成結構複雜的生物薄膜,由圓球形的酵母菌、假菌絲與菌絲等各種型態的細胞與胞外基質組成,其基因表現與游離的細胞有很大的不同,其中Efg1、Tec1、Bcr1、Ndt80、Brg1與Rob1等六個轉錄因子對生物薄膜的形成特別重要,另外可能有上千個基因都對生物薄膜的形成有所影響。生物薄膜可以多種機制提高白色念珠菌對抗真菌劑的抗性,包括增加外排泵的表現以將藥物排出胞外,以胞外基質中的水解酶分解藥物,以及形成抗性很強的休眠細胞(persister cells)。生物薄膜中的酵母菌還可以向外傳播,這些細胞比起正常的酵母菌型細胞有較強的黏附力,形成新的生物薄膜的能力較強,具有較高的致病能力。
白色念珠菌在環境中與人體中皆可形成生物薄膜。在醫療器材上,白色念珠菌是最常被發現的真菌種類,導尿管、靜脈導管、心律調節器、人工瓣膜、人工關節、隱形眼鏡與假牙等醫療器材上均可能有白色念珠菌的生物薄膜生長。生物薄膜中的白色念珠菌較正常游離者對人體免疫系統的抗性較高,可能藉由醫療器材入侵循環系統,造成嚴重的全身性感染,還可能使這些醫療器材無法正常運作。
在人體中,白色念珠菌還可能與許多細菌共同形成生物薄膜,包括口腔中的轉糖鏈球菌、格氏鏈球菌、粘性放线菌或細梭菌,腸道中的鬆脆桿菌、產氣莢膜梭菌、大腸桿菌、克雷伯氏肺炎菌與糞腸球菌,陰道中的乳桿菌屬細菌與囊腫纖維症患者肺中的綠膿桿菌,白色念珠菌與這些細菌可透過實際接觸、分泌訊息分子或改變環境因子來影響彼此的行為,例如綠膿桿菌可分泌一種內酯,抑制生物薄膜中白色念珠菌菌絲的形成,但不影響其整體生長速率。還有研究顯示白色念珠菌形成的生物薄膜提供了缺氧的微環境,使鬆脆桿菌與產氣莢膜梭菌等厭氧細菌得以生長,且這些細菌可能可以促進白色念珠菌形成生物薄膜,以利自己在有氧環境下生長。白色念珠菌與細菌共同形成的生物薄膜對人體健康的影響仍不清楚。
感染
白色念珠菌為伺機性感染的致病真菌,平時可生長於人體的皮膚與粘膜組織中而不造成負面影響,但當其過度生長時可能造成念珠菌症的感染。念珠菌症又可依感染部位分為若干類型,包括感染口部的鵝口瘡、感染陰道的念珠菌性外陰陰道炎、感染食道的念珠菌性食道炎等,甚至可能造成全身的侵入性感染,侵犯多個器官,並造成念珠菌血症(Candidemia)。念珠菌症在健康成人中比較少見,小於一個月的新生兒、愛滋病患者、癌症患者、糖尿病患者與服用抗生素與皮質類固醇等免疫缺乏者感染的風險較高。白色念珠菌也常造成醫療照顧相關感染。
淺表與局部感染
口腔感染

據統計,世界上有一半的人群口腔中有白色念珠菌,甚至有更精密的方法顯示90%的人皆有,但絕大多數不造成任何感染症狀,免疫缺乏、高糖份的飲食、配戴假牙、抽菸與口乾皆會造成口腔中白色念珠菌出現的機率增加。若白色念珠菌造成口腔粘膜損傷即為鵝口瘡,通常不伴隨其他症狀且沒有痛感,只有部分患者會有灼痛感,其他可能的症狀還包括味覺改變、觸感改變、患部發紅等。醫護人員常透過簡單觀察口腔即作出鵝口瘡的診斷,有時則需取小量檢體進行培養,再於顯微鏡下觀察鑑別。治療鵝口瘡使用的抗真菌劑包括克催瑪汝、咪康唑與耐絲菌素,較嚴重者則可能施以氟康唑。
食道感染
念珠菌性食道炎為白色念珠菌造成的食道感染,多發生於愛滋病患者、接受化療的癌症患者等免疫缺乏的人群,但有時亦見於免疫力正常者中。念珠菌性食道炎的症狀包括疼痛與吞嚥困難,其診斷多為使用內視鏡,治療多使用氟康唑。感染念珠菌性食道炎的患者通常同時患有口腔感染。
陰道感染

白色念珠菌為兩成女性陰道菌叢的正常成分,並不造成感染,若因激素、服用藥物或免疫力下降導致陰道環境改變,而使白色念珠菌大量增殖,便可能導致念珠菌性外陰陰道炎。免疫缺乏、服用避孕藥、服用抗生素與妊娠均會增加感染機會,症狀包括陰道痛癢、性交與排尿時有痛感與異常的分泌物,診斷多需取小量樣本進行培養,再於顯微鏡下觀察鑑別。治療方式於陰道塗抹抗真菌劑或口服氟康唑,嚴重者除繼續口服氟康唑外,還可能於患部施以硼酸、耐絲菌素或氟胞嘧啶。
念珠菌性外陰陰道炎是第二常見的陰道炎,僅次於細菌性陰道病。有高達75%的女性在其一生中得過至少一次念珠菌性外陰陰道炎,接近半數超過兩次。約5%的女性在一年中被感染三次以上。
其他
除了以上部位之外,白色念珠菌還可感染指甲而造成念珠菌性甲癬,感染甲溝造成念珠菌性甲溝炎,感染嘴角造成口角炎,感染舌頭導致正中菱形舌炎,感染男性生殖器導致龜頭炎,感染毛囊導致毛囊炎,在消化道過度生長而造成腸胃不適等。產道存有白色念珠菌的孕婦若發生早期羊膜囊破裂,可能造成新生兒感染先天性皮肤念珠菌病。
全身性感染
白色念珠菌亦可造成全身性的感染,侵入血液、心臟、眼、骨骼等部位,其中最常見者為念珠菌血症(Candidemia),為常見的醫療照顧相關感染,在美國每年約有46000例,患者多為原本就患有其他重大疾病者,高風險者包括裝置靜脈導管的患者、住院期間長的患者、加護病房的患者、施用廣效型抗生素的患者、燒燙傷患者、接受全靜脈營養治療者與嗜中性白血球低下者。
全身性念珠菌症通常是以血液樣本培養進行診斷,多數患者最初治療的推薦用藥為棘白菌素,感染的菌株種類與抗藥性確認後則改為氟康唑,部分患者也會以氟康唑為起始用藥。成功將血液中的白色念珠菌清除並消除症狀後,療程還需持續兩週。1999年,一篇發表於《Clinical Infectious Diseases》的研究統計美國49間醫院3年間所有在院內發生的血液感染,共記錄了超過一萬起感染,其中念珠菌屬真菌為第四常見的病原,且以白色念珠菌為大宗。念珠菌血症的死亡率可高達40%-60%,每年造成的醫療花費超過十億美元。
註腳
參考資料
外部連結
| 维基共享资源中相关的多媒体资源:白色念珠菌 |
- Candida Genome Database (页面存档备份,存于互联网档案馆)
- U.S. National Institutes of Health on the Candida albicans genome
- Mycobank上白色念珠菌的頁面 (页面存档备份,存于互联网档案馆)
- 研究念珠菌的實驗室列表 (页面存档备份,存于互联网档案馆)
- Protein-protein interactions for Candida albicans (页面存档备份,存于互联网档案馆)
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||