
遗传连锁(英語:genetic linkage,或称基因连锁)是指有性生殖中的减数分裂期在同一染色体上两个位置靠得很近的基因的DNA序列有同时被遗传的倾向。若两个遗传标记所在的基因座相互靠得很近,则在染色體互換过程中它们被分离进入不同染色单体中的可能性较小,并因此得名为遗传“连锁”。换言之,同染色体上两个基因的距离越近,二者发生基因重组的概率就越低,就越容易被同时遗传。不同染色体上的基因是完全不连锁的,但其可能有害的等位基因的外显度可能受其他等位基因的影响,而这些其他等位基因可能位于别的染色体上。
遗传连锁是孟德尔定律最明显的例外。首个证实存在遗传连锁的实验完成于1905年。当时,人们还不知道为什么某些性状会共同遗传。后来的研究揭示了基因是与物理距离相关的物理结构。
一般用厘摩(cM)单位来衡量遗传连锁的程度。两个遗传标记之间的距离为1 cM表示每100个减数分裂产物中,两者在1个中分离到了不同的染色体,或者说每50次减数分裂发生1次。
发现
孟德爾的自由组合定律提出,每种性状的继承是与其他性状相互独立的。但对他的实验进行重复时,研究者发现了例外情况。1905年,英国遗传学家威廉·贝特森、伊迪丝·丽贝卡·桑德斯和雷金纳德·庞尼特在实验中做了和孟德尔的做法相似的豌豆杂交实验。他们对甜豌豆的性状遗传有兴趣,并且正在研究两个基因——花色(P紫色和p红色)基因和花粉粒的形状(L长和l短)。他们杂交了纯种PPLL和ppll并对子代PpLl进行自交。
根据孟德尔定律,表型PL:Pl:pL:pl的比例应当为9:3:3:1。然而,他们观察到的是更高的PL和pl频率、更低的Pl和pL的频率:
| 表型及基因型 | 观测值 | 9:3:3:1理论值 |
|---|---|---|
| 紫,长(P_L_) | 284 | 216 |
| 紫,圆(P_ll) | 21 | 72 |
| 红,长(ppL_) | 21 | 72 |
| 红,圆(ppll) | 55 | 24 |
该实验揭示了P与L等位基因和p与l等位基因之间的连锁。 P与L同时出现和p与l同时出现的频率,大于重组的Pl和pL。F2杂交比回交更难计算重组的频率,但上表中实际观察到的重组子代数与期望的子代数不吻合,甚至不到50%。这表明两个因素以某种方式相互作用,阻止其他两种表型的出现,而产生了这种差异。他们得出了这样的结论:一些性状是相互关联的,因为它们在染色体上彼此接近。
托马斯·亨特·摩尔根的工作扩展了对连锁的理解。摩尔根观察到,连锁基因之间的交叉互换数量不同,这使得交叉频率可能表明染色体上基因分离的距离。以他的名字命名命名的单位“厘摩”(centimorgan)被用作表示交叉互换的频率。
连锁图谱
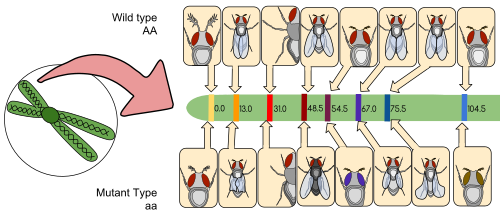
连锁图谱(也称遗传图谱)是一个物种或实验种群的表格,用于展示其已知基因或遗传标记之间的用重组频率推算出的相对位置,而非它们在每个染色体上的具体物理距离。连锁图谱是由托马斯·亨特·摩尔根的学生阿尔弗雷德·斯特蒂文特首先提出的。
连锁图谱是一种基于同源染色体交叉过程中,遗传标记间重组频率的图谱。两个遗传标记之间的重组(分离)频率越大,就认为它们相距越远。历史上最初使用的标记是由编码区DNA序列产生的可观测表型(酶的产生、眼睛的颜色);后来,研究者也使用已证实或假定的非編碼DNA序列,如微衛星或产生限制性片段長度多態性(RFLPs)的片段。
通过检测已知遗传标记的连锁,连锁图谱可以帮助研究人员定位其他遗传标记(例如其他基因)。在绘制连锁图谱的早期阶段,人们用这些资料来识别基因连锁群,即一组已知存在连锁的基因。随着知识的进步,更多的标记可以添加到一个连锁群中,直到这个连锁群覆盖整个染色体。对于已得到充分研究的生物体,其连锁群与染色体之间是一一对应关系。
參見
外部链接
- Genetic Mapping
- A list of computer programs for genetic analysis including linkage analysis
- Linkage versus Linkage-Disequilibrium
| |||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||