
脂肪酸合成是指利用乙醯輔酶A以及丙二醯輔酶A經過脂肪酸合酶的催化,反應合成脂肪酸的過程。這對細胞和生物體內的脂肪生成作用與糖解作用是相當重要的一項流程。該過程發生在細胞的細胞質中。 轉化為脂肪酸的大部分乙酰輔酶A通過糖酵解途徑來源於糖类。
直鏈脂肪酸
飽和直鏈脂肪酸

細胞通過不斷重複進行下表六個類似β氧化的反應可以獲得具有十六個碳的棕櫚酸。 左圖描繪了大腸桿菌中的脂肪酸合成酶II(縮寫為FASII,一個由多種酵素結合成的複合體)如何產生飽和直鏈脂肪酸,這種酵素複合體能夠在原核生物、真菌、植物、寄生蟲以及粒線體內找到。
大多數真菌、酵母菌以及動物體內的飽和直鏈脂肪酸是利用脂肪酸合成酶I進行合成。脂肪酸合成酶I是一種巨大而多功能的二聚體蛋白質酵素,具有催化脂肪酸合成過程所有步驟的能力。脂肪酸合成酶I不僅能夠合成直鏈脂肪酸也能夠合成具有支鏈的脂肪酸。
在合成出(16:0)棕櫚酸(16個碳,0個雙鍵)之後,可以再經由一連串的修飾延長碳鏈或是增加不飽和度(或兩者皆是)。延長的產物,如18:0硬脂酸。進行延長作用的地點是在內質網上,而延長的反應通常由一個蛋白質催化完成,但其複合體的物理結構可能可以協助其反應進行。
| 步驟 | 酶 | 反應 | 描述 |
|---|---|---|---|
| (a) | Acetyl CoA:ACP transacylase |
|
將乙醯輔酶A轉化為乙醯ACP |
| (b) | Malonyl CoA:ACP transacylase |
|
將丙二醯輔酶A轉化為丙二醯ACP |
| (c) | 3-ketoacyl-ACP synthetase |
 |
使乙醯ACP與丙二醯ACP反應,延長其中一個ACP |
| (d) | 3-ketoacyl-ACP reductase |
|
還原醯基部分C3上的那個酮基 |
| (e) | 3-Hydroxyacyl ACP dehydrase |
|
脫水 |
| (f) | Enoyl-ACP reductase |
|
去除C2=C3間的雙鍵(還原作用) |
上表中的ACP為醯基攜帶蛋白(Acyl carrier protein)
調控機制
乙醯輔酶A經過乙醯輔酶A羧化酶的作用可以轉化為丙二醯輔酶A,當反應進行到這一步時,便注定要完成脂肪酸合成這一條代謝路徑了(這稱為承諾反應,committed step)。而乙醯輔酶A羧化酶也同時受到異位調控以及磷酸化調控(磷酸化調控常見於哺乳動物)。其中乙醯輔酶A羧化酶的異位調控會受到棕櫚醯輔酶A(脂肪酸合成的最終產物)抑制,而被檸檬酸(在克氏循環中與乙醯輔酶A一起被合成出來)活化。生物可以藉由這點調控飽和直鏈脂肪酸的合成。
人類的從頭合成
De Novo在拉丁文中是「新」的意思,因為人類是從無到有合成脂肪酸,所以這樣命名。人類的脂肪酸主要是由肝臟以及哺乳期婦女的乳腺製造,也有一部分的脂肪酸是由脂肪組織合成。主要原料乙醯輔酶A大多是在粒線體中由丙酮酸脫氫酶分解丙酮酸而來,接著乙醯輔酶A與草醯乙酸被檸檬酸合成酶合成檸檬酸,再將檸檬酸移往細胞質中由ATP檸檬酸裂解酶再次分解成乙醯輔酶A和草醯乙酸。最後乙醯輔酶A將作為脂肪酸合成的原料,而草醯乙酸則被還原為蘋果酸返回檸檬酸循環中。
去飽和
去飽和指的是讓飽和脂肪酸轉為不飽和脂肪酸的過程,這過程需要氧分子、NADH以及細胞色素 b5的參與。脂肪酸的去飽和反應,發生在細胞的內質網上,會造成脂肪酸的氧化並生成一個NADH。在生物中的去飽和反應有無氧以及有氧兩種。而脂肪酸上最常發生去飽和的地方是在C9-C10的地方。
不飽和脂肪酸不論在真核生物或是原核生物中都扮演極重要的角色:細胞膜上主要的成分就是不飽和脂肪酸,因為細胞膜中的不飽和脂肪酸可以保持細胞膜的流動性,或著成為細胞膜上傳遞訊息、供其他細胞辨識的分子。
無氧去飽和
許多微生物用無氧的去飽和反應合成不飽和脂肪酸,在16:0棕櫚酸被合成後先去飽和形成雙鍵接著再進行延長的動作。

- 這整個反應由FabR以及FadR的轉錄調控控制。其中FadR蛋白扮演了兩個重要的角色:fabA與fabB序列轉錄的活化因子以及β-氧化的抑制因子。FabR蛋白則是fabA與fabB序列轉錄的抑制因子。
有氧去飽和

有氧去飽和是將飽和脂肪酸轉為不飽和脂肪酸最常見的方式,在所有的真核生物以及部份的原核生物中都可以觀察到。藉由這條反應途徑可以將飽和脂肪酸轉化為具有相同碳數的不飽和脂肪酸,也可以在脂肪酸的支鏈上加上雙鍵。在枯草桿菌(Bacillus subtilis)中具有可以在Δ5位置加上順式雙鍵(cis-double bond)的酵素 Δ5-Des,而在釀酒酵母菌(Saccharomyces cerevisiae)中則有在Δ9位置加上順式雙鍵的酵素 Ole1p。 釀酒酵母菌(S. cerevisiae)利用雙調控系統(Two-component regulatory system)控制去飽和反應,藉由DesK和DesR這兩種蛋白之間的互動來調控。其中DesK是一種位在細胞膜上的激酶而DesR則是des基因的轉錄調控因子。這套調控系統受到溫度影響,當溫度降低時會增加脂肪酸的產量;當溫度升高時脂肪酸產量則會下降。
銅綠假單胞菌
在正常情況下,無氧和有氧的不飽和脂肪酸合成不會在同一個系統內發生,但銅綠假單胞菌(Pseudomonas aeruginosa)和弧菌(Vibrio)ABE-1為例外。銅綠假單胞菌會同時進行一個無氧的去飽和反應以及兩個有氧的去飽和反應。其中一個反應是利用DesA蛋白催化生成用來構造細胞膜的不飽和脂肪酸。另一個反應則是利用DesB以及DesC兩種蛋白來進行脂肪酸輔酶A的不飽和化,這個反應受到DesT的抑制。而DesT同時也是fabAB序列轉錄的抑制因子。
非直鏈脂肪酸
具有支鏈的脂肪酸通常都是飽和脂肪酸並依型態分為兩個族群:iso-series以及anteiso-series。
α-酮酸合成路徑


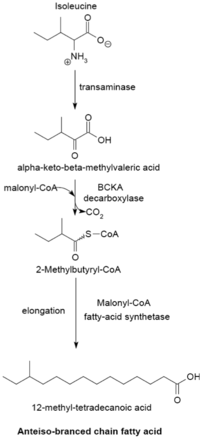
本系統以α-酮酸作為引子合成支鏈脂肪酸,見於大部分生物中(包含部分細菌)。α-酮酸系統的反應由纈氨酸、亮氨酸和異亮氨酸的轉氨化以及去羧化開始,經過這兩種反應合成3-甲基丁醯輔酶A和2-甲基丁醯輔酶A。由纈氨酸反應得到的2-甲基丁醯輔酶A可以作為引子進行延長,合成具有偶數碳數的iso-series脂肪酸。由亮氨酸反應得到的3-甲基丁醯輔酶A則可以作為引子進行延長,合成具有奇數碳數的iso-series脂肪酸。最後,由異亮氨酸反應得到的2-甲基丁醯輔酶A則可以作為引子進行延長,合成具有奇數碳數的anteiso-series脂肪酸。
若是藉由酵素BCKA(branched-chain α-keto acid decarboxylase)對這些引子前驅物進行去羧化,則可以經由與大腸桿菌合成飽和直鏈脂肪酸類似的路徑去合成支鏈脂肪酸。這條反應路徑的產物是一群碳數為12到17的脂肪酸。其脂肪酸種類惠依照細菌種類而定(不同的細菌會依此路徑合成不同脂肪酸),故也可以依此分類細菌。
BCKA脫羧酶與相對應的α-酮酸
BCKA是一個由兩個次單元(A2B2)所組成的四聚體酵素,為合成支鏈脂肪酸所必需的酵素之一。BCKA的功能主要是將α-酮酸去羧化,這項功能在支鏈脂肪酸的合成上遠比在直鏈脂肪酸的合成重要。而在芽孢桿菌屬生物中的重要性又特別高(芽孢桿菌屬由α-ketoisocaproate以及α-ketoisovalerate合成脂肪酸)。BCKA對支鏈α-酮酸的高親和力造就了其對於支鏈脂肪酸的合成不可或缺的地位。
| Substrate | BCKA activity | CO2 Produced (nmol/min mg) | Km (μM) | Vmax (nmol/min mg) |
|---|---|---|---|---|
| L-α-keto-β-methyl-valerate | 100% | 19.7 | <1 | 17.8 |
| α-Ketoisovalerate | 63% | 12.4 | <1 | 13.3 |
| α-Ketoisocaproate | 38% | 7.4 | <1 | 5.6 |
| Pyruvate | 25% | 4.9 | 51.1 | 15.2 |
影響碳鏈長度的因子
以α-酮酸作為引子合成支鏈脂肪酸時,產物通常都是碳數介於12到17之間的脂肪酸,其確切的碳數與物種有關。但是丙二醯輔酶A的濃度、溫度以及溫度穩定因子(Heat stable factor,HSF)的參與都會影響其結果。雖然上述因素皆可以影響產物的碳數,但溫度穩定因子的影響僅限於仰賴BCKA催化的反應途徑。 丙二醯輔酶A的濃度越高,產物中C17的脂肪酸比例就越高(直到C17脂肪酸的濃度到達大約20μM為止)。而桿菌生產C17脂肪酸的比率則依溫度下降些微降低。
短鏈分子合成路徑
這套系統以異戊酸、異丁酸和2-甲基丁酸這種短鏈分子作為引子合成支鏈脂肪酸,出現於無法使用α-酮酸作為引子的細菌-瘤胃菌(ruminal bacteria)中。其所需的材料(即異戊酸、異丁酸和2-甲基丁酸)在環境中皆可輕易取得,其總反應式為:
- Isobutyryl-CoA + 6 malonyl-CoA +12 NADPH + 12H+ → Isopalmitic acid + 6 CO2 12 NADP + 5 H2O + 7 CoA
此套系統與α-酮酸合成路徑最大的差別是其主要酵素的功能是將乙醯輔酶A轉換為乙醯-ACP。
ω-脂環族脂肪酸
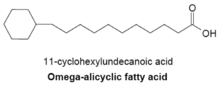
典型的ω-脂環族脂肪酸在ω端(甲基尾端)具有3碳或4碳環狀結構,在細菌中十分常見。合成ω-脂環族脂肪酸的酵素通常也能合成細胞膜上的支鏈脂肪酸,在一些細菌中甚至以ω-脂環族脂肪酸作為主要成分,因為環狀小分子引子在細胞中的供給量遠比直鏈狀引子多。 目前我們對這條反應的機制仍不清楚,但可以確定的是其途徑是由醣類轉化為莽草酸最後生成環已烷酸類輔酶A作為引子合成脂肪酸。
結核硬脂酸

結核硬脂酸(D-10-Methylstearic acid)是一種由一個分枝杆菌属(Mycobacterium spp.)以及兩個鏈黴菌屬(Streptomyces)的物種所製造的脂肪酸。以油酸(oleic acid)為前驅物與S-腺苷甲硫氨酸(S-adenosyl-methionine)進行酯化(esterify)形成一種磷脂,以NADPH輔助因子進行反應,最後產生10-甲基硬脂酸。
參考文獻
外部連結
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||

| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||