
| 生物分类学 |
|---|
| 分类阶元 |
|
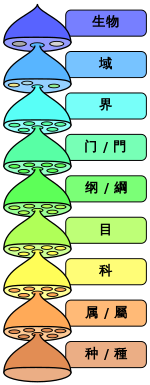
域 | 界 | 门 | 纲 目 | 科 | 属 | 种 |
| 二名法 |
| 三域系統 |
| 介於生物和非生物(無生物)之間的物體——不屬於任何一分類界 |
| 細胞生物 |
种(拉丁語:Species)或稱物种,是生物分类的基本单位。物种位于生物分类法中最底级,在属之下。籠統来讲,同一物种的任意两个性别适当的健康个体,都能够交配繁殖出具生殖能力的後代。满足以上条件的最大群体称为物种。根据演化生物學家恩斯特·麥爾的定义,物种是:「能够相互配育的自然种群的类群,且这些类群与其它的类群在生殖上相互隔离。」昆虫学家陈世骧(1978)对物种所下定义为:「物种是繁殖单元,由又连续又间断的居群所组成;物种是进化单元,是生物系统线上的基本环节,是分类的基本单元。」
在分类学中,一个物种被赋予一个拉丁化的雙名法名称。该名称使用斜体印刷,手写时则加上底線;属名首字母大写,屬名之後紧跟一个唯一的形容词,這個詞稱為種小名或種加詞,其首字母不可大寫。只有完整的双名制名称才称为「种名」,而非仅仅是双名制名称的第二个部分。例如人的种名叫Homo sapiens(智人),而不是sapiens。
物种也是演化和生物多样性的基本单元。 在19世纪,生物学家们意识到物种可以在足够的时间内进化。查尔斯-达尔文于1859年出版的《物种起源》一书中解释了物种如何通过自然选择而产生。这一认识在20世纪通过遗传学和种群生态学得到了极大的扩展。基因变异来自突变和重组,而生物本身是可移动的,由此导致地理隔离和遗传漂变两种不同的选择压力。基因有时可以通过水平基因转移在物种之间交换;新物种则可以通过杂交和多倍体迅速产生;物种也可能因各种原因而灭绝。然而,病毒是一种特殊情况,具有平衡的突变和选择的概率,可以被视为准种。
物种的定义
物种的定义:常是指一类形态和遗传组成相似的生物群体,有生殖能力的个体之间在自然状态下能产生正常后代。以下是其他方式的物种定義:
- 形态学种(morphological species)是一群在外观上不同的生物,例如我们可以分辨出鸡和鸭,因为它们长着不同形状的喙,而且鸭子的足有蹼。在有记录的历史之前,物种就已经像这样定义了。尽管受到不少批评,形态学种的概念仍然是日常生活中最常使用的物种概念,而且在生物学中依然扮演着重要角色,尤其是在植物学中,而且此概念對於無性生殖的生物體也可以適用。
- 生物学种(biological species)或隔离种(insular species)概念把物种定义为一组能够或潜在能够相互培育的生物。对于研究现存高等生物,如哺乳类、鱼类和鸟类的科学家来说,这不失为一种简洁的表达方式;但对于不进行有性生殖的生物来说,这个定义是毫无意义的。它把互相配育在理论上的可能性和种群间基因流动的可能性区分开来。例如马和驴杂交是可行的,并且能够产生后代,然而它们仍然属于两个种。这有两个原因:第一,因为在野生状态下,马和驴通常不会杂交。第二,因为它们结合产生的后代很少具有繁殖能力。定义生物学种的关键在于两个种群间没有显著的基因物质交流。值得注意的是:有的物種較早分化,但仍未產生完整的生殖隔離;有的物種較晚分化,但已經產生完整的生殖隔離。
- 配偶识别种(Mate-recognition species)定义了一群能够把种群中的其它个体识别为潜在配偶的生物。正如上述隔离种的概念那样,它仅适用于进行有性生殖的生物。
- 系统发生种(Phylogenetic species)或者进化种或者达尔文种是一组具有相同祖先的生物;在时间和空间上,这个世系与其它世系保持分离。在这样一组生物在的某个时候,其成员可能分道扬镳。当其不同越来越显著的时候,这两个种群就被认为是两个不同的种了。
在实际操作中,这些定义通常都相當一致,这些定义的不同点主要是侧重点不同而非相互否定。然而,还没有提出一个物种概念可以在无需具体分析情况下,就能完全应用于所有物種的。事實上,科學界對於物種的定義約有二十多種。物種問題就是指生物學家針對物種定義的爭議。
在生物学分类上的重要性
物种的概念历史悠久。这之所以是最重要的分类阶元之一,有下面的原因:
- 它是普通人区分不同生物的基础- 狗是一种,猫是另一种。
- 科学家通常使用标准的双名制命名法(或三名制命名法)来指代生物。
- 这是唯一的具有经验成分的分类阶元,比如断言两只动物是两个物种就不单单是对它们进行分类那么简单了。
在使用了几千年后,这个概念依然集中在生物学和相关学科上,也仍然被错误的定义和争论不休。
“指定物种状态”的含义
对于某个物种的命名应该看作是对于该群生物的进化关系及其独特性的一种假定。更新的资料会证实或推翻这个假设。有时候,尤其是旧时通讯不发达的时候,分居异地的分类学家会为后来定为同一种的生物命两个不同的名字。当两个经过命名的种发现其实是同一个种时,先发表的那个种名通常会保留,后发表的被废弃,这一过程称为synonymization,或lumping。将一个分类阶元分开成多个通常是新的阶元称为splitting。分类学家常被他们的同时戏称为“lumpers”或“splitters”,视乎他们找到了物种间的不同点还是相同点。(参阅統合派與分割派)。
传统上,研究人员依赖于解剖学上的不同,和通过观察不同种群间是否有成功混交来区分物种;解剖结构和生殖行为依然是指定物种状态的重要手段得益于上几个十年里微观生物学研究技术的日新月异,包括DNA分析等,得出了一大批关于物种间相似与区别的知识。许多从前认为是不同物种的类群现在归入了单一的分类阶元,许多过去归在一起的类群被分开了。任何的分类阶元(种,属,科等)都可以合并和分开。对于一个较高的阶元,这样的修订则更显得意义深远。
从分类学的角度看,种下的类群定义为低于种的分类阶元。在动物学中,仅使用亚种概念,而植物学还会使用诸如变种,亚变种,和变型等单位。
更详细的隔离种的概念
总的说来,对于那些大型复杂的有性生殖生物(例如哺乳类和鸟类), 生物学种或隔离种概念的定义有几种变体。通常,区分两个物种,甚至是亲缘关系很近的两个物种都是很简单的。例如,马(Equus caballus)和驴(Equus asinus)不需要经过专门训练或学习的人就能够把它们分开。然而它们相近得可以进行杂交,并产生不育的骡。它们明显就是不同的种。
但是许多情形则让人难以决定。这就是隔离种和进化种的分歧之处。两个概念都同意一个物种在时间上保持着其血统的独立性,它与其他这样的血统可以被我们区分(否则我们就认不出它们来了),是生殖隔离的(否则一个种群如果一有机会就和另外一个合并起来了),并有有效的种内识别机制(没有这个,该血统不可能延续)。在实际上,两种观点都同意一个种必须有其自身的独立进化历史;否则刚才提到的特征都不适用了。其不同在于:进化种概念不对种群的未来作出预测,而仅仅是记录已知的类群。相反,如果经过研究人员的研究,某两个种群在可能情况下能够再次合并,隔离种概念就不认为这两个种群为两个种。
隔离的问题
有两个关键的问题需要解决。第一,初定的这个种是否能持续的、可靠的与其它物种区分;第二,这在未来是否会持续下去。针对第二个问题,下面有几个地理分布上的可能性
- 初定的两个种分布区重叠的—它们占有相同的栖息地。经过多年的观察,都无法在重叠区分辨出有两个以上的种群,这些种群可能在随后时间又合为一个大种群。没有生殖隔离,种群就不得以分化,有了生殖隔离,种群间的基因无法交流,也就无法抹掉差异。这不是说杂交完全不会发生,只是说它可以忽略不计。通常,杂种个体无法像其亲本纯种个体那样成功的繁殖。
- 初定的两个种地理隔离—它们占有不同的地理区域。很明显,无法直接观察到分隔两地的两个种群之间的生殖隔离情况。通常情况下也不可能通过试验来完成:即使在豢养情况下两个初定的种能配育,这不代表在野外它们也会自由交配,这也不总是可以对杂种后代在进化上的适应性提供多少信息。通过其它一些试验可能会得出更多信息:例如,种群A的个体是否对种群B发出的求偶鸣叫录音有正确的反应呢?有时候,试验能够提供确切的答案。例如,巴拿馬地峽两边居住着七组看上去几乎一样的海虾(Altheus)种群,它们在约3百万年前出现。直到不久前,它们还被认为是相同的7个种。但是当把来自海峡两边的雄性和雌性放在一起时,它们没有交配反而大打出手。甚至如果海峡再次下沉,这些种群在基因上还会保持隔离:所以,它们现在被划分为不同的种。但在更多情况下,观察和试验都无法提供确定的答案,这是就要通过对邻近物种的经验来猜测它们之间的关系了。
- 初定的两个种地理比邻—它们的繁殖区域比邻但不重叠。这种情况很少有,尤其在温带地区。分界线通常都是栖息地突然改变的地方(群落交错区),比如像森林边缘或者山上的雪线等,但有时却不大起眼。地理上的比邻指出这两个种群占有很相似的生态位导致它们无法在同一个地区共存。又因为它们不进行杂交,可以肯定地说有一种机制(通常是行为上的)来防止种群间的基因交流,因此它们应该属于不同的种。
- 在两个种群混合区有杂交区域。典型情况下,杂交区域会存在'纯种'种群一方或双方的代表,加上第一代杂交种和反交得到的后代。通过对杂交区域的广度与存在疑问的两个种群典型散布区广度的比较,可以评估两个'纯种'种群之间基因交流阻力的大小。例如橡树的分布距离是鸟或松鼠携带橡子的距离; 袋食蚁兽的散布距离约为15公里,因为这是小袋食蚁兽在离巢后寻找空闲领地时通常会经过的距离。如果杂交区域相对于散布区域越窄,则两个种群间的基因交流就越少,它们之间就更倾向于在进化到路上继续分化。然而,很难去预测未来杂交区域的变化;无论把具有杂交的种群定为两个种还是一个种,都具有潜在的争议性。
- 种群的变化是渐变式的—在种群地理分布的两极,其典型个体间差异十分明显,但是在它们间的过渡是逐渐进行的。例如,北澳大利亚的树袋熊比南方的亲戚要小而且颜色也较浅,但是它们之间没有绝然的分界线:越向南部,树袋熊的个体就越大而且颜色越深;在中间地带的个体则体型和色泽都是中等。相反,在相同的地理范围内,黑背(北方)和白背(南方)钟鹊不从一种形态向另一种过渡;北方种群黑背,南方种群白背,在杂交区域“纯种”与杂合类型同样常见。树袋熊的变化是渐变式的(从北到南逐渐过渡,在小范围内的种群具有统一的外观),但钟鹊的变化不是渐变的。在这两个例子中,对其分类都存在一些不肯定,但是大多数的看法是两者的南北双方都不足以构成种。由于认为南北的钟鹊基因交流受到了限制,足以称它们为亚种(而非种);但树袋熊之间存在南北间实质的基因交流,所以专家甚至拒绝认定它们为两个亚种。
物種分類的歷史
在古時,人只是賦與一群類同的生物一個名詞。希臘哲學家,如亞里士多德,認為生物是可嚴格區分為不同的“物種”,而且物種是永恆不變的。
隨着接觸的生物愈多,人開始嘗試認真把生物分類。最初,人只是以外貌與習性分類,就像中國人把大多數水上生物稱作“魚”。
1686年時英國自然學家约翰·雷是第一次為物種一詞下生物學定義的人。而在十八世紀林奈首次以生殖器官作生物分類。他似乎只是認為這樣的分類有些意義,而並不是以為有近似生殖器官的物種有可聯繫。畢竟,當時的歐洲人相信創造論,即所有物種由神獨立創造,並無任何其他的聯繫。可能由於創造論,林奈對一些差異極微的物種感到困惑。他因而提出一些物種為完美的,樣板的;另一些則是仿造的。
及至十九世紀,多數自然學者開始明白物種在不斷地變异,而地球的古老也足以容纳長時間累積形成的巨變。因而,分類法開始強調物種之間是如何演變。拉马克認為生物可以把需要的特質遺傳給後代,例如長頸鹿就是因為吃愈來愈高的樹葉而長出愈來愈長的頸項。(這只是簡化的描述,拉马克的理論當然遠為深邃)
拉马克最大的貢獻在於提出不同物種之間可以有連續不斷的系谱。他在1809年的著作《动物学哲学》(Zoological Philosophy)是首個以邏輯理由否定創造論。當達爾文的進化論在1860年代被廣為接受後,卻掩蓋了拉马克的貢獻。及至二十世紀末,拉马克的理論重新獲得重視,成為适应突变的一個基礎。他另一曾被捨棄的目標為本進化論,亦發展成人工选择。 現代的分類學主要以遺傳序列為基礎。
物种的数量

據美国夏威夷大学和加拿大戴尔豪斯大学合作的的海洋生物普查科学家推測,地球上约有870万种物种(±130万),其中650万种物种在陆地上,220万种生活在海洋深处。
对于真核生物:
- 大约有777万种动物(其中953,434种已经得到了描述和分类)
- 大约有298,000种植物(其中215,644种已经得到了描述和分类)
- 大约611,000种真菌(霉菌、蘑菇,其中43,271种已经得到了描述和分类)
- 大约36,400种原生生物(其中8,118种已经得到了描述和分类)
- 大约27,500种色藻(其中13,033种已经得到了描述和分类)
以上统计不包括病毒。
參見
| ||||||||||||||||||||||||||||||||