
種族(Race),是在体质形态或遺傳特徵上具有某些共同特色的人群,划分时可以根據外在特征(如肤色、髮色、面部骨骼结构等)、基因、以及自我认同为标准。人種这一概念以及种族的具体划分都是具有相当争议性的课题,其在不同的时代和不同的文化中都有差异,种族的概念也牵涉到诸如社会认同感以及民族主义等其他范畴。人种不属于生物分类学,在智人以下没有所谓人种的生物分类。
20世纪以前,科学家普遍认为,人类分为若干个本质主义方式划分的(即以不可缺的特征来划分的)人種,如尼格罗人种(南撒哈拉非洲人)、高加索人种(西歐亞人)、蒙古人种(東歐亞人)、澳大利亞人種(澳大利亞和美拉尼西亞原住民)等。但自1940年代起,演化科学家开始淘汰这种理论。另外,种族长期以来理解为一种科学分类,即将种族视为一个分类的层次,如将种族等同于亚种;但1960年代起,群体遗传学研究中新出现的数据以及模型也使一些科学家开始质疑这种理解,而转而以群体(population)、特征线(cline,又譯「漸變群」或「生態群」)等其他概念来研究人类内部的差别。1990年代以来,基因体学以及分支系统学研究中新出现的数据和模型也使科学界对人类起源有了新的认识,使一些科学家转而用世系而非特征来定义种族的划分,并且认为种族应该理解为模糊集合,统计群体,或广义的民族。
有许多演化学家以及社会学家认为,基于近年来的生物学研究结果,任何对于人类种族的定义,都缺乏科学分类的严谨性和正确性;人種的定义是不准确的、随意的、政治化的和约定俗成的,随文化视角的差异而变化,种族应该视为一种社会建构。但也有其他科学家认为,人種概念并非毫无根据,人種概念和多位点基因数据之间的确有关联,而试图摒弃人種概念的真正动机,主要为政治原因而非科学。
目前,各个学科对于人種是什么,是否存在,到底有几个,应该如何定义,如何理解,如何分析等问题,尚无定论或共识。
| 概念 | 参考 | 定义 |
|---|---|---|
| 本质主义的 | Hooton (1926) | 人类的大分类之一,主要区分于其内部共有的某些特征的组合,而这些组合源于共同的祖先,组成一个模糊的生理背景,一般上被个体之间的差别所掩盖,最好以一个组合的图样来理解。 |
| 基于科学分类的 | 恩斯特·麥爾(1969) | 一个种之内,一些表现型相似的群体的组合,居住在该物种的分布地域的一个区域内,从科学分类角度和该物种的其他群体不同。 |
| 基于群体的 | 多布然斯基(1970) | 人種是基因上相异的孟德尔遗传群体。他们既非个体也非某个基因型,而是多个基因相异的个体。 |
| 基于世系的 | Templeton (1998) | 亚种(人種)是在一个物种之内,有别于其他进化世系的世系。这个定义要求,该亚种的基因差异化必须是由长期存在的,防止基因流动的障碍造成的;也就是说,亚种不但需要基因差异化,也需要历史延续性。 |
| 基于亲缘分支分类法的 | Levin (2002) | 人種意指以地理为基础的世系关系,主要以大陆或较大的地域为划分标准;人種指以大陆或次大陆划分的亲缘分支。在亲缘分支分类法中,基本的分类是世系单位,即祖先加上后代之支系,即按照现在分析所得出的种族。 |
人種概念的起源
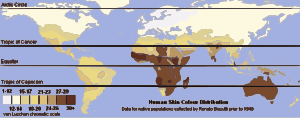
由于人类的成员擅长识别其他成员外观,其内部群体关系又极其复杂,因此自史前时代以来,人类就可能对人種的概念有一定的认知和理解,但各个文化对于人種的理解又不尽相同。最早记载种族的文学作品,为古埃及的《地狱之书》,其中将人类分为“埃及人”、“亚洲人”、“利比亚人”和“努比亚人”四类,可看出这个分类融合了人種、“民族”、“国家”等概念。后来的中国、罗马等文化比较注重于氏族,对于以外观划分的“民族”的认知则较少(Dikötter 1992;Goldenberg 2003)。而中国人在古代則往往认为华夏和四夷的人都是炎帝或者黄帝的子孙。不过古代中国后来也曾记载到白种人,一般称其为“黄毛人”,唐朝以后也记载了黑种人。希腊、罗马、中国等文化对外观不同的人種也有一定的认识,同时也导致其古代文献中(如中国的《山海经》)出现许多在遥远地方所存在的奇异种族的描写。同时也有一些罗马作家认为种族的特征是由居住环境决定的(Isaac 2004)。但在许多古代文明中,外表不同的人仍然可以通过采纳所在社会的文化标准而成为该社会完全的一员(Snowden 1983;Lewis 1990)。古代犹太人以及欧洲中世纪根据《圣经》认为人类是挪亚三子闪、含以及雅弗的后代,地球由三洲组成,其中闪族为亚洲人,含族为非洲人,雅弗族为欧洲人。
近现代对于种族的理解是欧洲地理大发现时代的产物(Smedley 1999)。欧洲人在探索世界的过程中,接触到了世界各地的许多民族,对于这些民族之间外表上的、行为上的、以及文化上的差异产生了许多猜想。同时由于非洲奴隶贸易使欧洲人的奴隶来源渐渐地由欧洲、中东转变成非洲,欧洲也因此产生了将人类分类的动力,以作为奴役并虐待非洲人的理由(Meltzer 1993)。
通过借鉴希腊罗马的文献以及当时欧洲内部的关系,例如英格兰人与爱尔兰人之间的不和(Takaki 1993),欧洲人开始将自己以及其他民族划分为外表、行为、能力皆有分别的群体,并把可以遗传的外表特征和内在的智力、行为、甚至道德水平互相联系(Banton 1977)。虽然其他文化也有相似的观点(Lewis 1990;Dikötter 1992),但是这种观点开始影响社会的构造,主要是在欧洲及其殖民地开始的。
传统的人種划分

对种族划分的最早的科学性尝试发生于17世纪,那正值欧洲帝国主义和殖民主义蓬勃发展的时期。最早出版的脱离罗马希腊传统的人类划分法是弗朗索瓦 ·貝爾尼埃(François Bernier)所著的Nouvelle division de la terre par les différents espèces ou races qui l'habitent(《基于地球上居住的不同的物种或种族的新的地球划分法》),于1684年出版。
Bernier共区分4个人種:
18世纪时,人类群体之间的区别成为科学研究的重点(Todorov 1993)。早期的学者注重于总结及描述“人类的自然类别”,这即是約翰·弗里德里希·布盧門巴赫(Johann Friedrich Blumenbach)于1775年出版的人类五分法的文章的标题。但随着19世纪时人类学的形成,欧美学者开始试图解释各个群体在行为上和文化上的不同特征(Stanton 1960)。他们开始测量颅骨的大小及形状,试图解释智力或者其他方面的不同特征(Lieberman 2001)。与达尔文于1859年出版《物种起源》同时,欧洲人对于不同种族到底起源相同,还是各自进化,或各自被上帝创造,展开了激烈的争论(Wolpoff and Caspari 1997)。
17世纪至19世纪期间,民间对于民族之间的区别的普遍认知,和科学家对于这些区别的解释结合在一起,形成了后来一名学者所说的“人種的意识形态”(Smedley 1999)。这种意识形态的基本要旨是:种族是远古的,自然的,一成不变的,各不相同的。虽然一些群体是多个群体的混合体,但是通过调查和研究,仍然可以辨别形成该混合群体的祖先民族。
19世纪时,有多位自然科学家在种族的问题上发表了自己的观点,如喬治·屈維埃(Georges Cuvier)、詹姆斯·考維斯·普里查德(James Cowles Pritchard)、路易·阿加西(Louis Agassiz)、查爾斯·皮克林(Charles Pickering)、約翰·弗里德里希·布盧門巴赫(Johann Friedrich Blumenbach)。其中屈維埃将人类三分,普里查德七分,阿加西八分,皮克林十一分。布盧門巴赫的五分法则是19世纪时比较常见的:
- 蒙古人种,即黄色人种,主要分布在中亚、东亚等地
- 衣索比亚人种,即黑色人种,主要分布在非洲、大洋洲等地
- 美洲人种,即红色人种,主要分布在美洲
- 马来人种,即棕色人种,主要分布在东南亚(注意:和20世纪时所说的棕色人种不同)
- 高加索人种,即白色人种,主要分布在欧洲、西亚等地
在布盧門巴赫之后的几十年里,研究人员渐渐将美洲、马来两个人种归并于蒙古人种,结果进入20世纪初剩下三个主要的人种:
20世纪最常见的分法是由美国人类学家卡爾頓·S·昆(Carleton S. Coon)提出的:
传统人種和其他人类现象之间的联系
19世纪的自然科学家对于种族的认识可以归纳为三点:
- 人種是客观存在的,自然发生的人类分类;
- 人種和其他人类现象(如行为、文化、智力、道德水平)等有很深的联系,因此也造成了不同文化之间物质文明丰富与否的区别;
- 人種因此是一个有效的科学分类,可以用来解释以及预测个人或群体的行为。
人种以肤色、眼色、眼型、髮色、髮質、鼻型、嘴唇的厚薄、头型、脸型等特征划分,而这些生理特征却和其他文化特征,甚至智力、道德水平联系了起来。如:当时认为,高加索人种的浅肤色和高眉骨显示了高加索人种高深的智力水平以及仁厚的心灵;蒙古人种的浅黄肤色和内眦赘皮显示了其狡猾,并且有些死板教条的性格;尼格罗人种的深肤色、低眉骨、厚嘴唇则显示他们比较接近于猿类动物。(但值得一提的是:黑猩猩的皮肤其实非常白,嘴唇也很细。)
进入20世纪以后,这种种族主义的观念,乃至“种族”的概念本身,都已经开始面临越来越多的挑战。
20世纪、21世纪时围绕人種概念展开的争论
人種研究的范围
由于对于种族的研究在至少两个范围(国家范围以及国际范围)进行,同时各种研究的目的也不同,因此对于种族的讨论也非常复杂。一般来说,进化论科学家将人类作为一个整体对待,在他们对全球人类多样性的研究过程当中,有层次的科学分类所提供的帮助有限,或者根本不适用。但是在国家层面运作的政府、执法人员、医药事业等则比较注重于在国家或者地区内所出现的基因多样性,对他们来说,有层次的科学分类的确颇有实用价值。
研究范围和目的的不同,从近年来发表的三篇研究论文中就可以看出:Rosenberg等(2002年)、Serre & Pääbo(2004年)、以及Tang等(2005年)。Rosenberg等以及Serre & Pääbo所研究的都是全球人类基因的多样性,但是他们的结论却完全不同,Serre & Pääbo将此不同归于实验设计。Rosenberg等从全球各地取样,他们没有把地理因素考虑在内,而Serre & Pääbo则是根据地理而取样。通过从各洲主要群体取样,Rosenberg等认为有证据表明统计群体的存在(即种族)。但Serre & Pääbo认为,从地理的角度来考虑,人类基因多样性是渐变式的,特征线式的。Rosenberg等的研究方向是医学(即流行病学),而Serre & Pääbo的则是人类进化。Tang等研究的是美国国内的基因多样性,主要方向是研究种族和地理位置之间哪一个对于流行病学研究影响更深。与Serre & Pääbo不同,Tang等认为人種更为重要。最近的研究也将人種和群体基因结构相联系,和Tang等同出一辙。但国际范围和国内范围的研究为何会得出的不同结论,Serre & Pääbo所推测的原因是:
人種和亚种
随着20世纪初现代综合论的兴起,生物学家发展出了新的,更严谨的,将种族等同于亚种的定义。对于这些生物学家来说,人種是一个种中,组成其全部或一部分的,可辨认的群体。一个“单型”的种就没有人種(或者可以说是只有一个种族包含了整个种)。以下情况的种都可视为单型种:
- 该种所有成员都非常相似,没有有意义的方法将其分类。
- 该种的成员有相当的多样性,但是有很高的随机性,从遗传角度来讲毫无意义。(许多植物物种就属于此类,因此许多园艺学家假如要保留某种特征(如花瓣颜色)的话,会避免利用种子繁殖,而会使用插枝等不改变原有基因的途径。)
- 该种的成员有相当的多样性,且多样性有一定的规律可循,但是各个群体之间没有明确的界限,而是渐变式地互相融合。这种特征线式的多样性,一般上表示,各个看似分开的群体之间,其实存在相当的基因流。假如一个物种的各群体之间有相当的基因流,该物种很有可能仍然是单型种。
一个多型种就是有两个或以上种族的物种。这种物种通常有多个明显的亚种,一般上不会杂交(但可能有较窄的杂交区域),但在有机会的情况下会进行杂交。(但注意:假如两个群体在有机会的情况下仍然不可杂交的话,那么这两个群体就属于两个不同的种,而非一种之内的两个种族。)
虽然这个以概念精确性为目的的尝试得到了许多生物学家的支持(尤其是动物学家),但是进化论科学家却从多个方面对其进行批评。
人種概念的摒弃以及“群体”和“特征线”概念的兴起
20世纪初,人类学家对于人種是有截然分别的语言、文化、社会群体的理论开始质疑,摒弃。之后,群体遗传学的兴起使一些在人类学和生物学等学科的主流进化论科学家开始质疑,人種是否是一个客观存在的科学概念。对种族概念持否定态度的科学家主要从四个角度分析:实证、定义、其他理论、道德伦理。(Lieberman and Byrne 1993)
首先从实证角度挑战种族概念的是两位人类学家:法蘭茲·鮑亞士(Franz Boas)展示了环境因素造成的表现型的可塑性(Boas 1912),Ashley Montagu则依靠基因学证据。(1941, 1942)动物学家Edward O. Wilson和W. Brown则从一般动物系统学的角度挑战该概念,同时否认了人種等同于“亚种”的理论。(Wilson and Brown 1953)
特征线概念的提出,是对人类表现型和基因型重新理解过程中最重要的发展之一。该概念源自人类学家C. Loring Brace所描述的现象:自然选择、迁徙以及基因漂变在塑造人类基因多样性的同时,其多样性主要呈渐变分布,而这些渐变中出现的渐变式变化曲线可称为特征线(Brace 1964)。这个现象突显出了以表现型(如肤色、髮質)来描述的种族忽略了许多其他的,和人種划分重合较少的特征(如血型)。因此,人类学家Frank Livingstone得出的结论是:因为特征线和种族界限互相交叉,因此根本没有所谓的人種,只有特征线。(Livingstone 1962: 279)1964年,生物学家保罗·埃尔利希及Holm指出了一些两个或以上条特征线以不协调地分布的特例:如黑色素的分布取决于离赤道的距离,越远就越少;但是beta-S血色素的单基因型却是以位于非洲的特定的地理位置为中心,呈放射性分布。(Ehrlich and Holm 1964)以人类学家Leonard Lieberman和Fatimah Linda Jackson的原话来说:
| “ | 人类基因异质性的不协调分布,也反证了任何把人类的群体描述成在基因型或甚至表现型方面同质性[的描述]。 | ” |
最后,基因学家Richard Lewontin鉴于人类多样性的85%皆出现于群体内而非群体之间的现象,提出论点,认为人種和亚种都不是正确的或有用的描述人类群体的方法。(Lewontin 1973,该论点后来被其反对者称为Lewontin的谬误。)有研究人员报告说,(以休厄尔·赖特之群体结构统计FST来测量的)种族之间的区别仅有人类基因多样性的5%。2但由于FST的技术局限性,许多基因学家认为很低的FST值并不反证人类种族存在的理论。(Edwards, 2003)同时,还有如大卫·哈维(1982, 1984, 1992)的新马克思主义者认为,种族是一个用来巩固阶级之间不平等的社会构造的手段,实际上并不存在。
这些从实证角度提出的对人種概念的挑战,迫使进化科学重新考虑“种族”的定义。20世纪中叶,人类学家William Boyd将种族定义为:
- 一个在其拥有的一个或多的基因的频率上,和其他群体有显著差别的群体。我们选择哪些基因位点,选择多少个,作为显著的“星座”,是完全随意性的。
Lieberman和Jackson(1994)指出:这个论点的弱点在于:假如一个基因就能区别人種,那么种族的数量就和进行交配的人类配偶数量一样多。同时,人类学家Stephen Molnar提出,特征线的不协调性不可避免地导致种族数量的大幅增加,最终使这个概念失去意义(Molnar 1992)。
在种种实证方面和概念方面的问题出现的同时,第二次世界大战之后,进化科学家和社会科学家很清楚地认识到,和种族有关的理念被屡次用来给种族歧视、种族隔离、奴隶制以及种族清洗提供理由。1960年代期间美国民权运动以及世界各地反殖民主义运动兴起期间,这个从伦理道德角度提出的质疑也日益激烈。
在这些论点的轮番攻击下,一些进化科学家完全摒弃了人種概念,转而使用“群体”。群体和种族的不同在于,群体所指的是一个繁殖群体(在进行遗传算法时必不可少的概念),而不是一个生物学上的分类。其他的进化科学家则采用了特征线的概念(即某个特征的频率相对于地理的变化速度)。“群体”和“特征线”的概念并不矛盾,许多进化科学家两者并用。
进化科学家摒弃人種概念的同时,许多社会科学家则把人種改换成民族,其中民族是指在国籍、宗教、等方面自我认同的群体。这些科学家完全承认,这些国籍、宗教、种族等方面的自我认同全部都是社会构造,和自然或超自然领域的客观事实可以完全无关。(Gordon 1964,另见:美国人类学协会在种族问题上的声明[2] (页面存档备份,存于互联网档案馆))
人类基因多样性的起源、模式以及外在体现
人类的起源

任何人类种族的模型都必须能解释人类进化过程中种族差异的形成。但在20世纪末以前,人类学家只能依靠相当不完整的化石记录来推断人类进化的过程。而他们的模型也无法作为我们就种族起源问题提出推论的有效基础。然而,近年来分子生物学领域的发展已经开始给进化学家提供更新、更全面的数据,也充实了我们对于人类起源的认识。
人类学家对于智人(homo sapiens;现代人)的起源,一向争论不休。大约一百万年前,直立人(Homo erectus)从非洲迁徙到欧洲和亚洲。而争论的中心就是:直立人是否是在非、欧、亚三洲各自同时进化为智人,还是智人在非洲进化之后,离开非洲并取代了欧亚二洲的直立人。两种模型的不同,也导致了各自对种族起源问题的解释上的不同。
多地起源说
支持多地起源说的学者(见Frayer et al. 1993)指出了中南欧洲(Smith 1982)以及东亚、澳洲(Wolpoff 1993)等地化学纪录中的解剖延续性,认为解剖延续性证明了基因亲和性。他们认为,人类内部显著的基因相似性并不表示他们有共同的祖先,而是反映了世界各地人类群体之间,互相联系所造成的不间断的基因流。(Thorne and Wolpoff 1992)他们同时认为这个模型和特征线的理论并不矛盾。(Wolpoff 1993)
这个模型在人種问题上最重要的元素是,既然智人在世界各地已经进化繁衍了一百万年,这个物种完全有时间进化成多个种族。然而,Leiberman and Jackson(1995年)却指出,这个模型取决于以下和人種有关的条件是否成立:
- 人属(Homo)在其分布地中心和边缘地的成员之间须在更新世中期就有显著的构成学区别;
- 许多特征须先在人属的分布地域的边缘处发展;
- 这些特征随时间推移并不消失。这种地域性的多样化即可作为人属内部长期以来即存在差别以及这些差别是现今人类内部差别的前身的证据。
非洲起源说
自1990年代起,多位点基因型普遍地用来区分不同的人类群体,并且用来将个体分类到群体中(Bamshad et al. 2004)。从这里得出的数据也使学者开始审视生物学上将人種定位为世系的可行性,以及以亲缘分支分类法描述人種的正确性。多位点基因型主要用来确定人类历史中的演化,通过这种方法得出的人種概念亦等同于广义上的世系。
人类基因多样性研究表明,非洲是所有现代人类的起源地;智人于14万至29万年前迁出非洲,取代了直立人(Cann等1987)。欧洲人、亚洲人和美洲原住民是一个单一的,互有关联的群体,是较迟从非洲迁出的,可分为东、西两支。
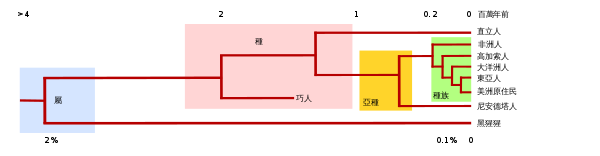 |
如上圖所示的基因進化樹一般由群體的脫氧核糖核酸(DNA)和蛋白質DNA排列得出,遠古人類血緣關係的研究主要通過粒線體DNA和Y染色體DNA排列進行,因為這些單一位點的DNA來源和一般的DNA不同,是從父母一方繼承的,不會經過重組合。各個以大陸為基準的群體內部的個體之間的相似程度,一般上會比不同群體的個體之間要高。進化樹的根源為人類和黑猩猩的,在非洲發源的共同祖先。橫向距離對應以下兩點:
- 基因上的相似度。人類和黑猩猩之間的基因差距約為2%,是現代人類內部差距的20倍。
- 時間上的距離。以粒線體分析為根據,人類的共同祖先存在於約20萬年前,而人類與黑猩猩的共同祖先存在於約400至700萬年前。黑猩猩和人类属于不同的属(genera),以青色表示,另外种、亚种、和人种的分类也有标示,其中人种仅为粗略标示。
 |
以上是以近鄰結合法畫出的,代表18個人類群體之間關係的系統樹。從圖中可以看出,歐洲、亞洲、大洋洲人、美洲人的祖先離開非洲後,高加索人種最先分出,然後是大洋洲人,最後分開的是東亞人和美洲原住民。

自1980年代起,研究亦表明,人类基因的多样性比其他的动物种要低。两个随机取样的人类个体之间的差距为约1000个核苷酸中出现一个,而黑猩猩则为500个核苷酸中出现一个。学者认为,这显示人类存在的时间相对来说较短,可能没有时间进化出不同的亚种。然而,人类基因共有约30亿个核苷酸,因此两个个体之间的差距约为300万个核苷酸。这些单核苷酸多态性(SNP)中的绝大多数都是中立的,但也有一些有确实的作用,影响人类之间表现型的差别。据估计,人类群体中约有1000万个SNP,其中较罕见的SNP的出现频率约为1%。
群體遺傳學的領域亦認為,中立多態性的分佈反映了人類各群體的歷史。學者認為,人類在經歷了一次人口瓶頸之後,以非洲為發源地,進行了迅速的遷徙和擴張,最後造成了10萬年前(約5000代前)的非洲和歐亞之間的分叉,以及4萬年前(約2000代前)歐洲和亞洲之間的分叉。
較小的群體的快速擴張有兩個主要影響。一、奠基者效應,新的群體的祖先僅攜帶了原先群體的基因多樣性的一部分,造成了基因上的區別;二、隨着不同群體之間的距離的拉大,不同群體之間發生交配的概率也隨之降低,這種選性交配減低了不同地理群體之間的基因流,拉大了群體之間基因差距。因此,早期的群體由於奠基者效應,造成較大的基因漂變;由於對基因流的限制,各個群體中新出現的多態性傳給其他群體的概率也相應降低。
這些新的數據,重新點燃了圍繞種族概念的爭論,主要爭論內容為新的數據的理解方式以及以數據得出的結論是否合理。大多數研究人員認為,以大陸為劃分基準的群體,並不能立為不同的亞種。然而亦有研究人員質疑,不同的進化世系是否應該以種族稱之。這些難點對於生物醫學尤其重要,因為患者自我定義的種族經常視為患者譜系的可靠指示。
將人種等同於世系:支持方
基因數據可用來推斷群體的結構,也可將個人分類到地理世系當中,該世系也經常和個人本身作出的自我認同相對應。
推斷群體結構時,首先須選擇出大量有信息價值的基因標記。此類研究通常表明,居住於同一個大陸上的人之間的相似程度比不同大陸上居住的人之間的相似程度要大。此類研究經常遭到批評,批評者認為群體定義是先天做出的。但即使群體定義的判斷僅以基因數據為基礎,群體結構仍可推斷出。例如Rosenberg等(2002)的研究通過使用377個基因標記,成功地將52個群體中的1056個個體分類為六個聚類,其中五個和主要地理區域相對應。
然而,在將個體分類的研究中,自我描述的種族並不能和譜系完全相對應。某些民族,如拉丁美洲裔民族,並沒有同質的譜系;非裔美國人具有西非以及歐洲的血緣,其中非洲血緣佔80%左右(Shriver等2003);歐裔美國人亦有歐洲和非洲的血緣,其中約30%的人的ba歐洲血緣在90%以下。綜上所述,人種可進而理解為按比例劃分的血緣。
無論如何,研究顯示,在美國國內,自我認同的種族和基因檔案之間存在非常準確地對應。Tang(2005)利用326個基因標記,在美國15個地點抽樣的3636個人當中,推斷出了4個基因聚類,並且正確地將所有的人分類到4個自我認同的種族當中(白裔、非裔、東亞裔、拉美裔),只有5個人出現錯誤(錯誤率為0.14%)。由此可以看出,美國人口的基因結構的主要決定因素為遠古譜系,和與之有密切關係的現代人種認同,而非現在的居住地點。
區分不同大陸血緣的基因的技術,也可用來區分大陸內部的不同血緣。然而,大陸內部的血緣研究應需要更多的基因標記,因為鄰近的群體的共同祖先應該較遲,基因的相似性較高,亦會出現較明顯的特徵線分佈。有人將特徵線的存在視為聚類不存在的根據(Kittles和Weiss 2003),也有人認為較低的差異使聚類的區分比較困難,但是聚類並非不能區分(Bamshad等2004)。也有研究持折衷觀點,認為人類基因多樣性既包括特徵線,也包括可示範的聚類。[3]
將人種等同於世系:反對方
Rachel Caspari(2003)認為,親緣分支按定義就是單系的(一個包括某祖先的所有後代的分類),而人種不是單系的,因此也不可能是親緣分類。
對於Lieberman、Jackson(1995)等人類學家來說,用親緣分類來證實人種,在方法論上和概念上都有嚴重的問題。他們認為,該模式的支持者在最初抽樣的分類中就已經使用了種族分類。例如,印度人、北非人和歐洲人被先驗地歸類為高加索人種,這本身就對概念的理解產生了限制和歪曲,掩蓋了其他世系關係,降低了由直接環境因素造成的基因差異的重要性,模糊了對於血緣關係的理解。他們還認為,重點應該放在特徵線的變化上,因為特徵線承認了適應性的遺傳差異,同時也強調了該差異並非以人種的形式集體存在。
最近另有研究顯示,之前常視為單一人種的地區,亦存在均勻的特徵線式的基因差異,而所謂的空隙則是抽樣造成的缺陷(Serre和Pääbo 2004)。這些學者並不否認親緣分支方面的研究的重要性,但是他們不認同對“人種”一詞的繼續使用,因為“群體”和“特徵線”已經足夠描述研究結果。
目前各學科對於人種的理解
圍繞着人種概念展開的辯論橫跨多個學科,在學界尚無共識。不僅如此,不同的研究對人種一詞的定義和運用也不盡相同,有用人種來指本質主義的分類、群體、親緣分支、單倍型類群的,也有乾脆不用人種一詞的研究。
19世紀時,種族是人類學的中心概念之一。1866年,倫敦人類學會的創始人詹姆斯·亨特(James Hunt)曾宣稱,人類學最重要的真理就是“人類不同種族之間明顯的心理和道德區別”。從20世紀起,這個思想已趨下風。
科學界對諸如“高加索”、“尼格羅”、“蒙古”人種等詞彙的支持,在近一百年裡有明顯的下降。這些詞彙原先指的是顱骨類型,源自顱面測量的技術,目前在法醫人類學等領域繼續使用。
從1932年起,介紹體質人類學的大學教科書開始摒棄種族的概念:1932年至1976年期間,32本書中有7本摒棄了種族的概念;1975年至1984年期間,33本中有13本摒棄種族;1985年至1993年期間,19本中有13摒棄了種族。1931年刊的《體質人類學雜誌》中有78%的文章採用人種或相似的詞語來描述生物學上人種分類的模式;1965年刊只有36%,1996年刊有28%。美國人類學協會目前的觀點為,人種的概念是社會及文化建構,種族的概念無法通過科學方法得到檢測或證實;人類的群體並非界限分明的群體,種族的概念對人類不適用。
有遺傳學家認為,人種不是有意義或啓發法價值的概念;群體之間的基因差異亦是無意義的,因為人種內部的差異比人種之間的差異要大,人種的特徵互相覆蓋,沒有明顯的界限。但也有遺傳學家認為,自我認同的人種、民族、地理祖籍等既有根據也有實用價值,這些分類和多位點基因數據中得出的聚類有對應關係,該對應關係也意味着,基因因素可能促成了某些群體之間目前無法解釋的表現型差異。
2001年2月,《小兒及青少年醫學檔案》(Archives of Pediatrics and Adolescent Medicine)請求文章撰寫者提不再以人種作為解釋變數,不再使用已過時的詞語;《新英格蘭醫學雜誌》(New England Journal of Medicine)、《美國公共衛生雜誌》等也有類似的動作。除此以外,美國國家健康學會宣佈,從2006年2月1日起,處理研究資金申請時將着重尋找研究者,在基層醫生當中研究並宣傳醫學上人種定性對於國家保健所產生的有害效果,並引用一名學術雜誌的編輯,稱“以人種和民族為根據的分析已經成為了分析上的本能(knee-jerk)反應”。
1985年進行的一次調查顯示(Lieberman等1992),當被問及“智人種中有生物學意義的人種”時,各領域的1200名科學家有以下比例的參與者對之持反對態度:
- 生物學:16%
- 發展心理學:36%
- 體質人類學:41%
- 文化人類學:53%
但該調查並沒有給出確切的定義,參與者可以以分類學或者群體的方式理解種族的概念。
社會對於人種概念的理解
在一般情況下,人種經常用來指民族,結果造成科學界和大眾對於人種一詞產生了理解上的差異。如在美國,“西班牙裔”(Hispanic)、“拉美裔”(Latino)等常被視為一個人種,但是也有人將拉美裔視為由不同民族背景共同組成的語言和文化群體(ethno-linguistic group);而歐洲人一般不會把南歐人排除在高加索人種之外,卻又常將按照舊時顱骨測量被劃分為高加索人種的羅姆人、印度雅利安人視為其他人種。
不同的社會中產生的人種建構
後哥倫布時代美洲的種族和民族多樣性
雖然人種的概念在大發現時代以後的許多社會的構造中扮演舉足輕重的角色,但該概念的不足之處也愈發明顯。在歐、亞、非洲,鄰近民族之間的特徵分佈是漸變性的;當時的人類學家約翰·布盧門巴赫(Johann Blumenbach)指出,“一種人類漸漸融入另一種,使你無法劃出他們之間的界限。”(Marks 1995, 54頁)但在美洲的一些地區,情況就完全不同了。新世界的移民大多來自舊世界的幾個相距較遠的地區:西北歐、西非、東亞、南歐。同時,移民人口開始互相融合,同時也和美洲原住民族進行融合;一項調查發現,美洲非裔有相當的歐洲血統:如牙買加人的調查抽樣有大約7%,而新奧爾良非裔居民的調查抽樣則有23%(Parra et al. 1998)。同樣地,由於不同人種之間的交融以及混血後裔和主體民族的混同,許多“歐裔”美洲人也有一定的非裔和原住民血統:在一次對於美國東北部某大學學生的調查中,約有30%自認為“白人”的對象有少於90%的歐洲血統(Shriver et al. 2003)。
在美國,各種社會上和法律上的常規的出現,使得混血人群漸漸地被歸類到簡單化的人種分類當中(Gossett 1997),而自1790年展開的十年一度的人口普查,也促使了人種分類的設立,並提供了將人們歸類到這些分類當中的動力(Nobles 2000)。如美國歷史上長期奉行“一滴血規則”,規定任何有一“滴”黑人血統的人都是黑人,直至1967年該規則的法律依據被美國最高法院推翻;此常規在美國社會雖然日趨式微,但仍遺留一定的影響,並且產生了只有部分黑人血統,但外表或許比較接近白人的人被視為黑人的現象,如美國眾議員G. K. Butterfield;歌手瑪麗亞·凱莉等。而在巴西等混血現象較為普遍的國家,和人種類似的社會分類則比較複雜、眾多、易變;人們基於社會地位、階級、血統、外貌等一系列特徵,時常在這些社會分類之間移動。(Mörner 1967)
美國的人種
2000年人口普查中的種族資料是由接受調查者自由填寫的,同時普查亦宣稱種族為“社會建構”,不應視為科學性的分類;普查中共有五個預設種族,即“白人”、“黑人或非裔美國人”、“美洲印第安人或阿拉斯加原住民”、“亞洲人”、“夏威夷原住民或其他太平洋島嶼民族”;接受調查者可以選擇多個人種,可以自行填寫其他民族名稱;另外,人口普查中沒有將“西語裔”(Hispanic)列成種族,而是另設問題來調查受訪者對“西語裔”的民族認同。總的來說,當今美國的人種界限,比美國歷史上森嚴的種族界限要寬鬆、模糊;與此同時,美國社會中存在明確的種族分類,這一點和拉丁美洲的一些國家(如巴西)很不一樣。
從美國建立至今,美洲原住民、非裔移民和歐裔移民都一直被分類到不同的人種當中。在北美開拓的前三個世紀當中,這三個人種的判斷標準較為相似,都以外貌、血統和社交圈子為標準。但從十九世紀末開始,三個人種的判斷標準之間出現了很大的不同。在南北戰爭結束後的重建時期,越來越多的美國人開始將任何有“一滴”黑人血統的人視為黑人。到二十世紀初期,這種“看不見的”黑人的概念在全國範圍內廣泛採用,並且寫進了一些州的法律。然而,美洲原住民的範圍則繼續以“血緣比例”(blood quantum)為標準;而白人則必須有“純正”的白人血統才可視為白人。
為了將混血情況相當普遍的美國人口分類,社會上出現了越來越多的種族類別,出現了“二分之一黑人”、“四分之一黑人”、“八分之一黑人”等極其細緻的分類;同時,按照這種標準得出的“種族”,往往異於本人所認同的人種。
美洲原住民和非裔美國人的評判標準之間的顯著差別,也使一些人類學家猜測其背後的政治原因。傑拉爾德·賽德(Gerald Sider)認為,這種規則的目的是集中並保護白人對權利、財富、土地的完全控制。由於大多數黑人長期以來從事農業勞動,“一滴血”規則因而保證了白人對農業勞動力的控制,保證黑人勞動力的供應源源不絕;而由於原住民則長期控制邊疆地區的大片土地,因此白人通過“血緣比例”的概念漸漸將原住民邊緣化,沖淡本來“純正”的原住民部落,並且佔據其土地。
而隨著來自拉丁美洲,主要說西班牙語的移民的增多,美國又出現了一個人種標籤,即所謂的“西語裔”(Hispanic)或“拉丁裔”(Latino)。實際上,拉丁美洲的民族構成和血統淵源非常複雜,有歐裔、非裔、原住民裔、各種混血裔等;還有和拉丁美洲無關的西班牙裔也被歸併到“西語裔”之內。總的來說,“西語裔”和其他種族類別之間有相當的重曡現象,美國人口普查中也把“西語裔”和種族類別分列開來,各自統計。
從1960年代起,“一滴血”規則日趨式微,漸漸退出美國社會;1967年美國最高法院判決該規則違反憲法,從此“一滴血”規則不再在美國法律中出現。但與此同時,黑人權利運動的擡頭,也使該規則漸漸被黑人運動所採納,以壯大黑人的人口及勢力。如美國政壇新秀,前參議員、2008年之新任美國總統巴拉克·歐巴馬的父親是黑人,母親是白人,本人一般被視為黑人;而2006年的一項調查顯示,當被告知歐巴馬的血統時,有55%的白人和61%的西語裔認為歐巴馬不是黑人,而是雙重人種,卻有66%的黑人仍然認為歐巴馬是黑人。
巴西的人種
和19世紀的美國相比,當代的巴西人口結構缺少明確的人種界限。在巴西,人種並不僅是靠血統決定的二種分類而已,而是由髮色、髮質、虹膜色、膚色等多種外在特徵所決定的若干種分類。這些分類之間沒有明確的界限,每個人的人種也不一定和父母或兄弟姐妹相同。
在一次調查中,調查者在巴西巴伊亞州的某漁村給一百名受訪者看三個姐妹的照片,並請求他們給這三名姐妹分類。在調查結果中,只有六組將三人分到同一個人種分類當中;有十四組將三人分到三個不同的人種分類當中。在另一次調查當中,一百名受訪者看了九張照片,卻給出了四十種不同的人種類別。除此以外,巴西人對各種分類的確切定義也沒有統一的說法:在一次調查中,40%的受訪者認為moreno claro(淺膚色深髮色歐裔)比mulato claro(淺膚色歐非混血裔)要“淺”,而60%的受訪者則剛好相反。除此以外,同一個人的人種的分類亦可隨時間、對象、場合等改變。巴西的人口普查中以受訪者本人的種族認同為基準,普查結果中有幾百種類別,其中包括“藍色”(比黑色要黑)和“綠色”(比白色要白)等類別。
巴西社會中也存在種族歧視:歐洲殖民主義的影響,使偏黑的人比偏白的人更容易受到歧視。因此,巴西的人種狀況並未達到“後種族、後種族主義”的階段。
人種概念和其他領域之間的關係
人種和種族主義
從啟蒙時代開始,種族主義的概念被用作奴隸制度的根據,“低等”民族理所當然地成為“高等”民族奴役的對象。在這種環境下,種族之間的差異被誇大成近似於物種之間的差異,對於弱勢民族的虐待因此被社會所接受。
亞瑟·德·戈比諾的《人種不平等之探源》(1853年至1855年)和瓦謝·德·拉普熱之“人類社會學”是19世紀時期種族主義理論歷史的里程碑。他們提出了在高加索人種以下的種族(如雅利安人種),將雅利安人種塑造成高貴和文明的化身,宣揚並且誇大雅利安人對人類文明的貢獻。戈比諾認為,世界上所有的文明,包括古埃及、中國、墨西哥、秘魯文明都是由雅利安人所建立的,同時只有白人才是人類始祖亞當的後代。19世紀末至20世紀初,種族主義在歐美達到最高點,最終造成了納粹德國在第二次世界大戰時期對猶太人和羅姆人系統性的滅絕屠殺。
二戰結束之後,種族主義在西方社會式微,各國紛紛取消種族歧視、種族隔離法案,並且立法保護民族平等。除此以外,許多國家亦採取平權措施,給予弱勢族群各種優惠,以縮小族群之間的差距;但反對者亦認為,平權措施是反向的種族歧視。
人種和智力
關於不同人種之間是否有差異的問題,不同的研究者提出了許多不同的觀點。有研究者認為,智商的遺傳因素很大,不同種族之間的智商差距多少反映了不同種族之間存在先天性的智力差別;也有研究者認為,文化差異、環境差異、種族歧視、語言障礙才是不同人種之間智商差異的主因;有研究者認為,智商並不能很好地反映智力,智力有很多種;有研究者指出,事實上世界各國的平均智商近年來以每十年3%的速度逐年遞增,即所謂的弗林效應,其變化速度大大超越任何先天因素可能經歷的變化,或許和營養的進步、教育的發展等環境因素有關;還有研究者認為,人種本身就是無意義的概念,因此“人種和智力”之間的關係也是無意義的。
参见
人種
只列現時中文對國際上不同人群的外表習慣劃分。
外部链接
| 查看维基词典中的词条「人種」。 |
- Features of Evolution and Expansion of Modern Humans, Inferred from Genomewide Microsatellite Markers - Lev A. Zhivotovsky,et al.
- Genetic variation, classification and ‘race’ - Lynn B Jorde,et al.
- Categorization of humans in biomedical research: genes, race and disease - Neil Risch,et al.
- Genetic Variation Among World Populations:. Inferences From 100 Alu Insertion Polymorphisms (页面存档备份,存于互联网档案馆) - W. Scott Watkins,et al.
- Population genomics a bridge from evolutionary history to genetic medicine - L.B. Jorde,et al.
- Microsatellite evolution in modern humans: a comparison of two data sets from the same populations - L. JIN,et al.
- Short tandem repeat polymorphism evolution in humans (页面存档备份,存于互联网档案馆) - F Calafell,et al.
- Human Population Genetic Structure. and Diversity Inferred from Polymorphic. L1 (LINE-1) and Alu Insertions (页面存档备份,存于互联网档案馆) - D.J. Witherspoon,et al.
- Reconstruction of Human Evolutionary Tree Using Polymorphic Autosomal Microsatellites - Qasim Ayub,et al.
- Implications of biogeography of human populations for 'race' and medicine (页面存档备份,存于互联网档案馆) - Sarah A Tishkoff & Kenneth K Kidd
- Using mitochondrial and nuclear DNA markers to reconstruct human evolution - Lynn B. Jorde,et al.
- Traces of Human Migrations in Helicobacter pylori Populations - Daniel Falush,et al.
- Understanding Human DNA Sequence Variation (页面存档备份,存于互联网档案馆) - K. K. KIDD,et al.
- Assessing DNA Sequence Variations in Human ESTs in a Phylogenetic Context Using High-Density Oligonucleotide Arrays (页面存档备份,存于互联网档案馆) - Jian-Bing Fan, et al.
- Large-scale SNP analysis reveals clustered and continuous patterns of human genetic variation - Mark D. Shriver,et al.
| |||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||
|