
| 免疫球蛋白样受体 | |
|---|---|
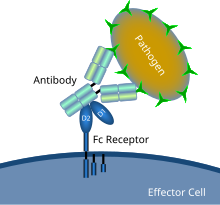 示意图显示了Fc受体与抗体包被的微生物病原体的相互作用
| |
| 鑑定 | |
| 標誌 | Fc受体 |
| 膜蛋白數據庫 | 10 |
Fc受体(英文:Fc Receptor)是在某些细胞表面发现的蛋白质,其中包括B淋巴球、滤泡树突细胞、自然杀伤细胞、巨噬细胞、中性粒细胞、嗜酸性粒细胞、嗜碱性粒细胞、人类血小板和肥大细胞——它们都有助于免疫系统的保护功能。它的名字来源于它对被称为Fc(可结晶片段区域)的抗体的一部分的结合特异性。Fc受体与附着在受感染细胞或入侵的病原体上的抗体结合。它们的活性通过抗体介导的吞噬作用或抗体依赖的细胞介导的细胞毒性刺激吞噬细胞或细胞毒性细胞破坏微生物或感染细胞。一些病毒(如黄病毒)使用Fc受体来帮助它们感染细胞,这种机制被称为抗体依赖性增强感染。
分类
有几种不同类型的Fc受体,它们根据它们识别的抗体类型进行分类。与最常见的一类抗体IgG结合的称为Fcγ受体,与IgA结合的称为Fcα受体,与IgE结合的称为Fcε受体。Fc受体的类别还通过表达它们的细胞(巨噬细胞、粒细胞、自然杀伤细胞、T细胞和B细胞)和每个受体的信号传导特性来区分。
Fcγ受体
所有的Fcγ受体都属于免疫球蛋白超家族,是调理素(标记)微生物吞噬作用最重要的Fc受体。该家族包括几个成员,Fcγ受体I(CD64)、Fcγ受体IIA(CD32)、Fcγ受体IIB(CD32)、Fcγ受体IIIA(CD16a)、Fcγ受体IIIB(CD16b),由于其不同的分子结构,它们的抗体亲和力不同。例如,Fcγ受体I与IgG的结合比Fcγ受体II或Fcγ受体III更强。Fcγ受体I还具有由三个免疫球蛋白样结构域组成的细胞外部分,比Fcγ受体II或Fcγ受体III多一个结构域。这种特性允许Fcγ受体I结合单个IgG分子(或单体),但所有Fcγ受体必须结合免疫复合物中的多个IgG分子才能被激活。
不同的Fcγ受体对IgG的亲和力各不相同,同样的,不同的IgG亚类对每种Fcγ受体都有独特的亲和力。这些相互作用由IgG CH2-84.4位置的聚糖(寡糖)进一步调节。 例如,通过产生位阻,含有CH2-84.4聚糖的岩藻糖会降低IgG对Fcγ受体IIIA的亲和力。相反,缺乏半乳糖并以GlcNAc部分终止的G0聚糖对Fcγ受体IIIA的亲和力会增加。
另一种Fc受体在多种细胞类型上表达,在结构上与MHC I类分子相似。该受体也结合IgG并参与该抗体的保存。然而,由于这种Fc受体还参与将IgG从母亲通过胎盘转移到胎儿或通过乳汁转移到哺乳期婴儿,因此它被称为新生儿Fc受体(FcRn)。最近,研究表明该受体在IgG血清水平的稳态中发挥作用。
Fcα受体
只有一种Fc受体属于Fcα受体亚组,称为Fcα受体I(CD89)。Fcα受体I存在于中性粒细胞、嗜酸性粒细胞、单核细胞、一些巨噬细胞(包括库普弗细胞)和一些树突状细胞的表面。它由两个细胞外Ig样结构域组成,是免疫球蛋白超家族和多链免疫识别受体(MIRR)家族的成员。它通过与两条Fc受体γ信号链结合发出信号。另一种受体也可以结合IgA,尽管它对另一种称为IgM的抗体具有更高的亲和力。这种受体称为Fcα/μ受体(Fcα/μR),是一种I型跨膜蛋白。这种Fc受体在其细胞外部分有一个Ig样结构域,它也是免疫球蛋白超家族的一员。
Fcε受体
已知有两种类型的Fcε受体:
- 高亲和力受体Fcε受体I是免疫球蛋白超家族的成员(它有两个 Ig 样结构域)。Fcε受体I存在于表皮朗格汉斯细胞、嗜酸性粒细胞、肥大细胞和嗜碱性粒细胞中。由于其细胞分布,该受体在控制过敏反应中起主要作用。Fcε受体I也在抗原呈递细胞上表达,并控制促炎性细胞因子的重要免疫介质的产生。
- 低亲和力受体Fcε受体II(CD23)是一种C-型凝集素。Fcε受体II作为膜结合或可溶性受体具备多种功能:它能控制B细胞的生长和分化,并阻断嗜酸性粒细胞、单核细胞和嗜碱性粒细胞的IgE结合。
汇总表
| 受体名称 | 主要抗体配体 | 对配体的亲和力 | 细胞分布 | 与抗体结合后的效果 |
| Fcγ受体I(CD64) | IgG1和IgG3 | 高(Kd~10−9M) | 巨噬细胞中性粒细胞嗜酸性粒细胞树突状细胞 | 吞噬作用
细胞活化 激活呼吸爆发 诱导杀灭微生物 |
| Fcγ受体IIA(CD32) | IgG | 低(Kd>10−7M) | 巨噬细胞 | 吞噬作用
脱颗粒(嗜酸性粒细胞) |
| Fcγ受体IIB1(CD32) | IgG | 低(Kd>10−7M) | B细胞肥大细胞 | 无吞噬作用
抑制细胞活性 |
| Fcγ受体IIB2(CD32) | IgG | 低(Kd>10−7M) | 巨噬细胞
中性粒细胞 嗜酸性粒细胞 |
吞噬作用
抑制细胞活性 |
| Fcγ受体IIIA(CD16) | IgG | 低(Kd>10−6M) | 自然杀伤细胞巨噬细胞(某些组织) | 诱导抗体依赖的细胞介导的细胞毒性作用(ADCC)
巨噬细胞诱导细胞因子释放 |
| Fcγ受体IIIB(CD16) | IgG | 低(Kd>10−6M) | 嗜酸性粒细胞
巨噬细胞 中性粒细胞 肥大细胞 滤泡树突细胞 |
诱导杀灭微生物 |
| Fcε受体I | IgE | 高(Kd~10−10M) | 肥大细胞
嗜酸性粒细胞 嗜碱性粒细胞 朗格汉斯细胞 单核细胞 |
脱颗粒
吞噬作用 |
| Fcε受体II(CD23) | IgE | 低(Kd>10−7M) | B细胞
嗜酸性粒细胞 朗格汉斯细胞 |
有可能的粘附分子
IgE跨肠上皮细胞转运 增强过敏性致敏的正反馈机制(B细胞) |
| Fcα受体I(CD89) | IgA | 低(Kd>10−6M) | 单核细胞
巨噬细胞 中性粒细胞 嗜酸性粒细胞 |
吞噬作用
诱导杀灭微生物 |
| Fcα/μ受体(CD351) | IgA和IgM | IgM为高,IgA为中等 | B细胞
系膜细胞 巨噬细胞 |
胞吞作用诱导杀灭微生物 |
| Fcμ受体 | IgM | 未知 | 人类Fcμ受体主要由淋巴细胞表达,但不由吞噬细胞表达 | 功能尚未完全阐明/多样化 |
| 新生儿Fc受体 | IgG | 在酸性细胞内体,高
在pH中性细胞外环境,低 |
单核细胞 | 通过胎盘将IgG从母亲转移到胎儿
通过乳汁将IgG从母亲转移到婴儿 保护IgG免于降解 跨内皮/上皮层转移IgG |
作用
Fc受体存在于免疫系统的许多细胞上,包括巨噬细胞和单核细胞等吞噬细胞、中性粒细胞和嗜酸性粒细胞等粒细胞,以及先天免疫系统(自然杀伤细胞)或适应性免疫系统(B细胞)的淋巴细胞。它们允许这些细胞与附着在微生物或微生物感染细胞表面的抗体结合,并帮助这些细胞识别和消除微生物病原体。Fc受体在其Fc区(或尾部)结合抗体,这种相互作用可激活拥有Fc受体的细胞。
激活吞噬细胞是Fc受体最常见的功能。例如,巨噬细胞在与Fcγ受体结合后开始通过吞噬作用摄取并杀死IgG包被的病原体。另一个涉及Fc受体的过程称为抗体依赖的细胞介导的细胞毒性作用(ADCC)。在ADCC期间,自然杀伤细胞表面的Fcγ受体III刺激自然杀伤细胞从其颗粒体中释放细胞毒性分子以杀死抗体覆盖的细胞。
Fcε受体I具有不一样的功能。Fcε受体I是粒细胞上的Fc受体,参与过敏反应和抵御寄生虫感染。当存在合适的过敏抗原或寄生虫时,至少两个IgE分子及其Fc受体在粒细胞表面的交联将触发细胞从其颗粒体中快速释放预先形成的介质。
信号机制 - Fcγ受体
激活
Fcγ受体属于免疫受体,它们共享相似的信号通路(涉及酪氨酸残基磷酸化)。这些受体通过称为免疫受体酪氨酸活化基序(ITAM)的重要激活基序在其细胞内产生信号。ITAM是一种特定的氨基酸序列(YXXL),在受体的细胞内尾部连续出现两次。当磷酸盐基团通过Src家族激酶的膜锚定酶添加到ITAM的酪氨酸(Y)残基时,细胞内会产生信号级联。这种磷酸化反应通常发生在Fc受体与其配体相互作用之后。ITAM存在于Fcγ受体IIA的细胞内尾部,其磷酸化会诱导巨噬细胞的吞噬作用。Fcγ受体I和Fcγ受体IIIA没有ITAM,但可以通过与另一种具有ITAM的蛋白质相互作用将并激活信号传递给它们的吞噬细胞。这种衔接蛋白称为Fcγ亚基,与Fcγ受体IIA一样,包含ITAM特有的两个YXXL序列。
抑制
仅存在一个YXXL基序不足以激活细胞,它代表一种基序(I/VXXYXXL),称为基于免疫受体酪氨酸的抑制基序(ITIM)。Fcγ受体IIB1和Fcγ受体IIB2具有ITIM序列,是抑制性Fc受体(它们不诱导吞噬作用)。这些受体的抑制作用受从酪氨酸残基上去除磷酸基团的酶控制,磷酸酶PTPN6和INPP5D抑制Fcγ受体的信号传导。配体与Fcγ受体IIB的结合导致ITAM基序的酪氨酸磷酸化。这种修饰产生了磷酸酶的结合位点,一个SH2识别域。ITAM激活信号的取消是由Src家族蛋白酪氨酸激酶的抑制引起的,并且通过水解膜PIP3中断激活受体的进一步下游信号,例如激活Fcγ受体s、TCR、BCR 和细胞因子受体(例如c-Kit).
Fcγ受体IIB的负信号传导对于活化B细胞的调节非常重要。阳性B细胞信号传导是由外来抗原与表面免疫球蛋白的结合启动的。分泌相同的抗原特异性抗体,它可以反馈抑制或促进负信号传导。这种负向信号由Fcγ受体IIB提供。使用B细胞缺失突变体和显性失活酶的实验已经证明了含有SH2结构域的肌醇5-磷酸酶(SHIP)在负信号传导中的重要作用。通过SHIP的负信号似乎通过SH2结构域与Grb2和Shc的竞争抑制Ras通路,并且可能涉及细胞内脂质介质的消耗,这些介质充当变构酶激活剂或促进细胞外Ca2+的进入。
细胞激活
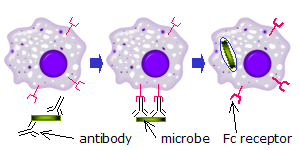
吞噬细胞
当对某种抗原或表面成分具有特异性的IgG分子通过其Fab(抗原结合片段区)与病原体结合时,其Fc区指向外,可直接到达吞噬细胞。吞噬细胞将这些Fc区与其Fc受体结合。许多低亲和力的相互作用在受体和抗体之间形成,它们共同作用以紧密结合抗体包被的微生物。低个体亲和力阻止Fc受体在没有抗原的情况下结合抗体,因此减少了在没有感染的情况下免疫细胞激活的机会。当没有抗原时,这也可以防止抗体对吞噬细胞的凝集。结合病原体后,抗体的Fc区与吞噬细胞的Fc受体之间的相互作用导致吞噬作用的启动。病原体通过涉及结合和释放Fc区或Fc受体复合物的活跃过程被吞噬细胞吞没,直到吞噬细胞的细胞膜完全包围病原体。
自然杀伤细胞
自然杀伤细胞上的Fc受体识别与病原体感染的靶细胞表面的IgG结合,称为CD16或Fcγ受体III。IgG对Fcγ受体III的激活导致细胞因子(如II型干扰素)的释放,这些细胞因子向其他免疫细胞发出信号,而释放出的细胞毒性介质(如穿孔素和颗粒酶)会进入靶细胞并通过触发细胞凋亡促进细胞死亡。自然杀伤细胞上的Fcγ受体III也可以与单体IgG(即未与抗原结合的IgG)结合。发生这种情况时,Fc受体会抑制自然杀伤细胞的活性。
肥大细胞

IgE抗体与过敏原的抗原结合。这些过敏原结合的IgE分子与肥大细胞表面的Fcε受体进行相互作用。与Fcε受体I结合后,肥大细胞的激活会导致脱颗粒,肥大细胞由此从其细胞质颗粒中释放预制分子(包括组胺、蛋白聚糖和丝氨酸蛋白酶在内的化合物的混合物)。活化的肥大细胞还会合成和分泌脂质衍生介质(如前列腺素、白三烯和血小板活化因子)和细胞因子(如白细胞介素-1、白细胞介素-3、白细胞介素-4、白细胞介素-5、白细胞介素-6、白细胞介素-13、肿瘤坏死因子、GM-CSF和几种趋化因子。)这些介质通过吸引其他白细胞而导致炎症。
嗜酸性粒细胞
曼氏血吸虫等大型寄生虫体型太大,无法被吞噬细胞吞噬。它们还有一个称为珠被的外部结构,可以抵抗巨噬细胞和肥大细胞释放的物质的攻击。然而,这些寄生虫可以被IgE包裹并被嗜酸性粒细胞表面的Fcε受体II识别。活化的嗜酸性粒细胞释放预先形成的介质,如主要碱性蛋白和酶(过氧化物酶),蠕虫对其不具有抗性。Fcε受体II与蠕虫结合IgE的Fc部分的相互作用会导致嗜酸性粒细胞以类似于ADCC期间自然杀伤细胞的机制释放这些分子。
T淋巴细胞
CD4+ T细胞(辅助性T细胞)为产生抗体的B细胞提供帮助。在疾病病理学中观察到几个激活的效应CD4+ T细胞亚群。Sanders和Lynch在1993年总结的早期研究表明,Fc受体在CD4+ T细胞介导的免疫反应中起着关键作用,并提出在细胞表面的Fc受体和T细胞受体之间形成联合信号复合物。Chauhan及其同事报告了标记的免疫复合物(IC)与活化的CD4+ T细胞表面上的CD3复合物共定位,因此表明Fc受体与T细胞受体复合物共存。观察到这两种受体在活化的CD4+ T细胞膜上形成顶端结构,这表明这些受体的横向运动。在细胞表面观察到Fc受体与T细胞受体和B细胞受体复合物的共迁移,T/B细胞偶联物在接触点显示这种共存。较早的报告表明Fc受体在CD4+ T细胞上的表达是一个悬而未决的问题。这确立了T细胞不表达Fc受体的当前范例,并且这些发现从未受到挑战和实验测试。Chauhan及其同事展示IC与激活的CD4+ T细胞的Fc受体配体结合。CD16a表达在活化的人类未成熟CD4+ T细胞中被诱导,这些细胞表达CD25、CD69和CD98,并且与IC的连接导致效应记忆细胞的产生。CD16a信号由Syk(pSyk)的磷酸化介导。
现在的一项研究表明,激活人类CD4+ T细胞后会诱导CD32a的表达,类似于CD16a。HIV-1研究人员的三项独立研究也表明了CD4+ T细胞上的CD32a表达。CD16a和CD32a在活化的CD4+ T细胞亚群中的表达现已得到证实。细胞表面的Fc受体在与由核酸组成的IC结合后触发细胞因子的产生并上调核酸传感途径。Fc受体存在于细胞表面和胞质溶胶中。CD16a信号上调核酸感应Toll样受体的表达,并将它们重新定位到细胞表面。CD16a是人类CD4+ T细胞的一种新的共刺激信号,它成功地替代了自身免疫过程中的CD28需求。在自身免疫背景下,CD4+ T细胞绕过CD28协同信号转导的要求而完全激活。此外,CD28协同信号传导的阻断不会抑制TFH细胞的发育,TFH细胞是产生自身抗体的自身反应性浆B细胞的关键亚群。免疫稳态需要共刺激信号和抑制信号之间的平衡。过度的共刺激或不充分的共抑制会导致耐受性崩溃和自身免疫。CD16a介导的共刺激在活化的CD4+ T细胞中提供阳性信号,而不是在缺乏Fcγ受体表达的静止细胞中。
参见
拓展阅读
-
Janeway CA, Travers P, Waldport M, Shlomchik MJ. Chapter 9. The Humoral Immune Response. Immunobiology: The Immune System in Health and Disease
 5th. New York: Garland. 2001. ISBN 978-0-8153-3642-6.
5th. New York: Garland. 2001. ISBN 978-0-8153-3642-6.
- Abbas AK, Lichtman AH, Pillai S. Chapter 12: Effector Mechanisms of Humoral Immunity. Cellular and molecular immunology 7th. Philadelphia: Elsevier/Saunders. 2012. ISBN 978-1-4377-1528-6.
- Gerber JS, Mosser DM. Stimulatory and inhibitory signals originating from the macrophage Fcgamma receptors. Microbes and Infection. February 2001, 3 (2): 131–9. PMID 11251299. doi:10.1016/s1286-4579(00)01360-5.
-
Maverakis E, Kim K, Shimoda M, Gershwin ME, Patel F, Wilken R, Raychaudhuri S, Ruhaak LR, Lebrilla CB. Glycans in the immune system and The Altered Glycan Theory of Autoimmunity: a critical review. Journal of Autoimmunity. February 2015, 57: 1–13. PMC 4340844
 . PMID 25578468. doi:10.1016/j.jaut.2014.12.002.
. PMID 25578468. doi:10.1016/j.jaut.2014.12.002.