
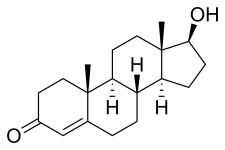
睾酮(主要的雄性激素)
| |
| 臨床資料 | |
|---|---|
| 其他名稱 | 雄激素、男性激素 |
| ATC碼 | |

雄激素(英語:androgen, androgenic hormones 或 testoids),亦作雄性激素、男性激素、男性荷爾蒙,是一类能够與雄激素受体(androgen receptor)結合,调节脊椎动物雄性性徵的发育与維持的類固醇激素的泛稱。雄性激素可以在睾丸、卵巢与肾上腺中天然合成,現代化學家也已發展出人工合成的方法。
男性体内的主要雄激素是睾酮,双氢睾酮(DHT)和雄烯二酮在男性的发育过程中同等重要。在胎儿时期,DHT导致了阴茎、阴囊和前列腺的分化;而在成年时期,DHT则会导致秃顶、前列腺增生和皮脂腺分泌旺盛。
虽然雄性激素常被认为仅是男性的性激素,但女性亦会产生雄性激素,只是数量较低,负责性欲与性唤起。雄性激素也是男女体内雌性激素的前体。
除自然作用之外,雄性激素亦可作为药物使用,如雄激素替代疗法和促进肌肉生长。值得注意的是,成年男性無論施打多少劑量的雄激素都無法讓生殖器再次發育成長,反而容易因劑量掌控不當而衍生出更多的生殖疾病。
类型
雄性激素以肾上腺雄激素(由肾上腺皮质最内层的网状带合成的19碳类固醇)为主,大多作为弱类固醇(weak steroids)产生作用(亦有部分用作前体)。
除睾酮之外,雄性激素还包括(于1970年代通过考虑所有生物学测定方法确定):
- 脱氢表雄酮(DHEA):由胆固醇转化而来,是多种雄性激素和雌性激素的主要前体。
- 硫酸脱氢表雄酮(DHEA-S):为 DHEA 的3β-硫酸酯和代謝產物,其在血液循环中浓度较大,但激素效应不大。
- 雄烯二酮(A4):睾丸、卵巢、肾上腺皮质均可生成,在代谢中被转化为睾酮和其他雄性激素,也是雌激素的母体结构。A4由于可被用作兴奋剂,已被国际奥林匹克委员会和其他运动组织禁止。
- 雄烯二醇(A5):类固醇的代谢产物,被认为是促性腺激素分泌的主要调节剂。
- 雄甾酮:雄激素分解过程中的副产物,也可以源于黄体酮。其在男性与女性的血浆和尿液中浓度大致相等,有轻微的雄性化作用,但效果仅为睾酮的七分之一。
- 双氢睾酮(DHT):睾酮的代谢产物,与雄激素受体的结合能力更强,作用比睾酮更为强效。在皮肤与生殖组织中合成。
生理功能
雄性激素的生理功能主要包括男性性器官在胚胎中的发育和男性第二性征在青春期时的发育。
男性胚胎发育
睾丸的形成
在哺乳动物的胚胎发育伊始,性腺即可成为卵巢也可成为睾丸。
对人类胚胎而言,大约第四周时,性腺的雏形出现于中间中胚层,与发育中的肾脏毗邻;大约第六周时,上皮性索在形成中的睾丸内发育,并在生殖细胞迁入性腺时与之合并。男性的某些Y染色体基因(尤其是SRY)控制男性表型的发育,包括将早期双能性腺转化为睾丸。男性体内的性索完全侵入发育中的性腺。
雄激素的生成
在发育中的睾丸里的性索中,来自中胚层的上皮细胞成为塞尔托利氏细胞,辅助精子的形成。
到人类胚胎发育的第八周时,少量的非上皮细胞(莱代细胞)会出现在肾小管之间,其在分化后不久就会开始产生雄激素。
雄激素的影响
雄激素是塞尔托利氏细胞所必须的旁分泌激素,也是男性胎儿显现男性特征所需要的(包括阴茎和阴囊的形成)。在雄激素的作用下,中肾及中肾管,发育为附睾、精囊和输精管。此效果来自雄激素与塞尔托利氏细胞分泌的抗穆氏管荷尔(MIH)的协同作用,MIH可以阻止胚胎的米勒管发育为输卵管和其他女性生殖组织。雄激素和MIH共同作用使睾丸进入阴囊。
早期调节
在约第11~12周垂体开始合成黄体生成素(LH)之前,人绒毛膜促性腺激素(hCG)会在第八周时促进莱代细胞的分化和其雄激素的生成。雄性激素在靶组织中的作用常涉及睾酮向 5α-双氢睾酮(DHT)的转化。
男性青春期
青春期时,男性与女性的雄激素水平均会上升,但男性雄激素水平的上升更为显著。雄激素介导男性第二性征的发展、生育能力的激活以及行为改变(例如倾慕女性或男性和性欲增强)。
男性的第二性征包括雄激素性毛发、声音变粗、喉结出现、肩膀变宽、肌肉增加和阴茎生长。
精子发生
在青春期,雄激素、黄体生成素和卵泡刺激素(FSH)分泌增加,性索空出,形成生精小管,生殖细胞开始分化为精子。在整个成年期,雄激素和 FSH 共同作用于睾丸中的支持细胞,以支持精子的产生。
外源性雄激素补充剂可作为男性避孕药。使用雄激素补充剂引起的雄激素水平升高可抑制LH的产生并阻断Leydig细胞产生内源性雄激素。
由于睾丸间质细胞分泌雄激素,如果睾丸中没有局部高水平的雄激素,生精小管就会退化,导致不孕。因此,许多经皮雄激素贴片被应用于阴囊。
脂肪沉积
男性的体脂通常比女性的要少。最近的研究结果表明,雄激素通过阻断一条信号转导途径来抑制某些脂肪细胞储存脂质的能力。
此外,雄激素(而不是雌激素)能够抑制α-2受体负反馈,从而在增加β肾上腺素能受体浓度的同时,降低α肾上腺素能受体浓度,提高作用于脂解诱导β受体的肾上腺素/去甲肾上腺素水平,进而减少脂肪的积累。
肌肉质量
相较女性而言,男性通常拥有更多的骨骼肌。雄激素能够促进骨骼肌细胞的增大,它可能通过作用于骨骼肌组织中的几种细胞类型(其中一种类型将激素信号传递给即成肌细胞),以协调的方式发挥作用。
雄激素水平升高会促进雄激素受体的表达,而成肌细胞融合产生肌管的过程则与雄激素受体水平密切相关。
脑
雄激素水平会影响人的行为,因为一些神经元对类固醇激素敏感,并且雄激素水平与人的攻击性和性欲调节有关。事实上,雄激素能够改变多个物种的大脑结构,包括小鼠、大鼠和灵长类动物,产生性别差异。
许多报告表明单靠雄激素就可以改变大脑结构,但由于两种性激素存在彼此转化的潜力,很难确定哪些神经解剖学上的改变是由雄激素或雌激素引起的。
来自雄性大鼠神经发生(新神经元形成)研究的证据表明,在确定雄激素对行为的影响时,海马体是一个有用的大脑区域。为了检测神经发生,对野生型雄性大鼠与具有睾丸女性化突变(TMF,一种导致对雄激素完全或部分不敏感以及缺乏外部雄性生殖器的遗传疾病)的雄性大鼠注射溴脱氧尿苷(BrdU),结果表明:
- 睾酮和双氢睾酮可以调节成年雄性大鼠的海马体神经发生(AHN)。
- 在野生型雄性大鼠中,海马体的神经发生是通过雄激素受体调节的,而在 TMF 雄性大鼠中则不是。
为了进一步测试活化雄激素受体对 AHN 的作用,对正常雄性大鼠注射氟他胺和双氢睾酮,发现 BrdU 标记细胞的增殖可由双氢睾酮促进,或由氟他胺抑制。
另有研究围绕轻度运动如何影响雄激素合成,进而导致 NMDA 的 AHN 激活展开,因为 NMDA 诱导钙流量,允许突触可塑性,这对 AHN 至关重要。一项研究向去势雄性大鼠和假去势雄性大鼠注射 BrdU,发现通过促进海马体中双氢睾酮的合成,轻度运动可以增加雄性大鼠的 AHN,但是 AHN 并不能通过激活雌激素受体而促进。
调节雄激素的水平亦可降低男性抑郁的可能性。一项针对青春期前雄性大鼠开展的研究表明,使用氟他胺治疗的新生大鼠与对照组大鼠相比,出现了更多的抑郁样症状。
社会隔离对 AHN 有阻碍作用,而雄激素的正常调节会促进 AHN。一项使用雄性大鼠的研究表明,睾丸激素可能阻碍社会隔离,导致 AHN 达到稳态;同期的 BrdU 分析表明,过量的睾丸激素并没有增加这种对社会隔离的阻断作用,亦即雄激素的自然循环水平抵消了社会隔离对 AHN 的负面影响。
女性特有效应
雄激素通过非基因组、雄激素受体独立的途径在松弛子宫肌层方面具有潜在作用,可预防妊娠期子宫过早收缩。
生物活性
雄激素结合并激活雄激素受体,以调节其大部分生物学效应。
| 激素类型 | 睾酮 | DHT | 雄烯二醇 | 雄烯二酮 | 脱氢表雄酮 | 雄甾酮 |
|---|---|---|---|---|---|---|
| 相对活性 (%) | 40 | 100 | 0.0008 | 0.04 | 0.02 | 0.06 |
DHT 在维持正常前列腺重量和导管腔质量方面的效力是睾酮的2.4倍,在防止去势后前列腺细胞死亡方面与睾酮具有同等效力。
雄激素也被发现通过膜雄激素受体发出信号,这与经典的核雄激素受体不同。
女性合成的雄性激素
卵巢与肾上腺的雄激素合成能力远低于睾丸。一项针对六名经期女性的研究观察了卵巢和肾上腺对女性雄激素水平的相对贡献,并得到如下结果:
- 在整个月经周期中,肾上腺对外周血中 T、DHT、A、DHEA 和 DHEA-S 的贡献相对恒定。
- 卵巢对外周血中 T、A 和 DHEA-S 的贡献在月经中期达到最大水平,而对外周血 DHT 和 DHEA 的贡献似乎不受月经周期影响。
- 卵巢和肾上腺对外周血中 T、DHT 和 A 有同等贡献,除了在月经中期卵巢对外周血 A 的贡献是肾上腺的两倍。
- 肾上腺提供了外周血中 80% 的 DHEA 和 90% 的 DHEA-S。
| 激素类型 | DHEA | DHEA-S | 雄烯二酮 | 睾酮 | DHT |
|---|---|---|---|---|---|
| 卵巢 (%) (F / M / L) |
20 | 4 / 10 / 4 | 45 / 70 / 60 | 33 / 60 / 33 | 50 |
| 肾上腺 (%) | 80 | 90~96 | 30~55 | 40~66 | 50 |
| F:卵泡早期,M:中期,L:黄体晚期 | |||||
参见
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||