

最後共同祖先(英語:last universal ancestor,縮寫為 LUA),又譯最近共同祖先,也被稱為最近普適共同祖先(last universal common ancestor,縮寫為 LUCA)、共通祖先(cenancestor)、原生命(progenote),演化生物學推導出來的假設,指地球生物最原始的共同祖先,是地球上所有現存生命的共同起源,但這共通祖先未必是最早的生命。學者一般相信,最後共同祖先在古太古代出現,距今約35億至38億年。最後共同祖先分化出細菌與古菌,演化成各種生命。
歷史背景

早在1809年拉馬克就在他的《動物哲學》中提及了生物演化樹;而查爾斯·達爾文在1859年於他的書《物種起源》提出的通用進化理論,並說:「所以我可以類推出,所有曾經生活在這個地球上的有機生物,大概都是從一種原始的形式中降生出來的,從第一個有呼吸的生命開始。」而英語中指稱此種原始生命的Last Universal Common Ancestor一詞,則首次見於帕特里克·福泰爾在1999年發表的一篇論文中。
在《物種起源》一書中,達爾文兩次提到他認為所有的生命都有共同的起源,他在結語中說:「所以我可以類推出,所有曾經生活在這個地球上的有機生物,大概都是從一種原始的形式中降生出來的,從第一個有呼吸的生命開始。」而他在書本的結語中重述了這個猜想:「這種生物起源的觀點有其宏偉之處,數種力量被注入了一種或數種生命形式之中。」而之後在1924年亞歷山大·伊萬諾維奇·奧巴林進一步提出了生命起源自由某種能量來源啟動的化學反應的理論。
所有生命的最後共同祖先可能擁有的特徵


盡管這共同祖先的大致解剖結構有很多不確定處,但根據地球上所有獨立生存的生物都具有的「普世」性質,還是可以以一定程度的細節描述其生物化學機制。
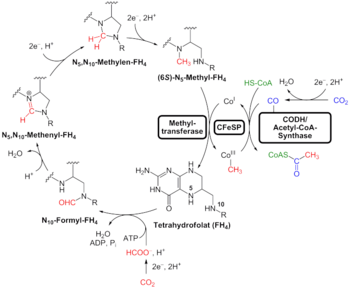
在2016年,瑪德琳·C·魏斯(Madeline C. Weiss)與其同事在對存在於源自己定序的、在系統發生樹上分屬不同位置的多種原核生物的大約六百一十萬個蛋白質編碼基因及286,514組蛋白質群集進行檢驗後,他們發現說其中有大約355組蛋白質群集很有可能存在於最後共同祖先當中,而他們的結果顯示說「最後共同祖先很可能是種厭氧、行固氮作用的嗜熱生物。這種生物很可能使用仰賴氫並固定二氧化碳的伍德-隆達爾代謝途徑(還原性乙醯輔酶A途徑);而最後共同祖先的生物化學反應中充滿了硫化鐵群集與自由基反應機制。」而其輔因子則「仰賴過渡金屬、黃素、辅酶A、鐵氧還蛋白、鉬蝶呤、咕啉和硒等等物質;而其遺傳編碼則需要核苷修飾和仰賴S-腺苷甲硫氨酸的甲基化作用等。」他們的研究產生了一個相對特定的結果,那就是產甲烷的梭菌屬細菌在355個檢驗過的生物類群中位於基群的位置;而最後共同祖先可能生活在位於地質活動區的無氧海底熱泉附近,而這樣的環境富含氫、二氧化碳與鐵。
基本可以肯定最後共同祖先有基因和遺傳編碼,而其基因很可能以DNA構成,也因此最後共同祖先出現於RNA世界之後;而在其有DNA的狀況下,其DNA編碼使用跟今日相同的四種核苷酸,也就是腺嘌呤(A)、鸟嘌呤(G)、胞嘧啶(C)和胸腺嘧啶(T);其DNA以DNA聚合酶這種模板依賴的酵素保持雙螺旋結構,而其DNA聚合酶可能屬於D族聚合酶;而其DNA結構的完整性,則仰賴於一類包含DNA拓撲異構酶在內的維護和修理DNA結構的酵素;此外,若其遺傳編碼基礎是DNA的話,那這些編碼會透過單股RNA作為中介來表現,而其RNa會透過仰賴DNA的RNA聚合酶生成,而其RNa的編碼會使用與DNA相似的核甘酸,但其中DNA的胸腺嘧啶會以尿嘧啶取代;另外這種生物會有多種包含組織蛋白摺疊蛋白在內的DNA結合蛋白。
這種生物的遺傳編碼會以蛋白質表現,而這生物體會用信使RNA(mRNA)轉譯DNA訊息,並透過核糖體、轉運RNA(tRNA)及一系列相關蛋白組成的機制,將自由胺基酸給組成這些蛋白質。其核糖體會由兩個子單位構成,其中大的單位是50S核糖體亞基,而小的單位是30S核糖體亞基,而每個子單位都會包含一個由核糖體蛋白包圍的核糖體RNA;而其核糖體RNA跟轉運RNA在核糖體的催化過程中都會扮演重要的角色。這種生物的蛋白質只會用上20種胺基酸,且只會使用左旋胺基酸,其他無數種的胺基酸都不會使用;另外,這種生物會使用ATP作為能量中介,且這種生物會使用數百種蛋白質酵素反應,以從脂肪、醣類和胺基酸中萃取能量,並以多種化學途徑合成脂肪、醣類、胺基酸跟核酸等。
這種生物會有著被磷脂雙分子層有效包含的、以水份為主的細胞質,這細胞會傾向使用特定的離子通道(或離子幫浦)排除鈉並聚集鉀;這生物的細胞會透過複製其內所有的物質及隨後發生的細胞分裂來繁衍。這細胞會利用化學滲透產生能量,並藉由乙酰跟硫酯還原二氧化碳及氧化氫分子(也就是產生甲烷跟醋酸鹽的過程)。
最後共同祖先很可能生活於類似海底熱泉的高溫環境中,而海底熱泉是因為海水與海床下的岩漿互動而產生的;而對最後共同祖先的後代的系統發生概括分析顯示,這最後共同祖先應當是一種微小且單細胞的生物,這種生物可能有環狀、自由漂浮在細胞內的DNA;而在形態學上,這種生物與小型現代細菌之間的分野可能不太明顯;而根據三域系統地提出者卡爾·烏斯的說法,最後共同祖先「大概是一種比三個域各自的祖先還要來得簡單、原始的生物。」
另一個研究「普世」特質的方法是利用基因分析以找出系統發生上古老的基因,而這樣給出的圖像是最後共同祖先生活於地球化學上嚴峻的環境且類似現代的原核生物;而對生物化學途徑的分析與系統發生學分析指向同樣的化學反應。實驗顯示諸如甲酸鹽、甲醇、乙醯類物質和丙酮酸鹽等乙醯輔酶A途徑的化學物質會在有水、二氧化碳及自然金屬的環境中自然產生,就如海底熱泉的環境一般。
年代
在2000年至2018年間,研究不斷地將最後共同祖先的生存年代往前推移,在2000年,研究認為最後共同祖先生活的年代是距今35億年前至38億年前的古太古代,也就是生命最早的化石證據的數百萬年前,而各候選證據的年代大約為距今34億8千萬年前至42億8千萬年前左右;而一篇在2018年來自布里斯托大學、使用分子鐘模型的研究,認為最後共同祖先的生存年代距今大約45億年前左右,也就是在冥古宙的時候。一般認為最後共同祖先(以及更一般的生命起源)不能存在於月球形成之前,,而這是因為根據大碰撞說,月球形成時地表會因融化與蒸發等原因而變得無法居住之故。
假說
在學界首次提出最後共同祖先的概念時,根據各活細胞間遺傳距離製成的分支圖認為古菌很早就從其他生物中分化出來,這點可從當時已知的古菌大多生活在諸如高鹽、高溫或高酸度等的極端環境推斷,而這也使得一些科學家認為最後共同祖先生活於類似今日海底熱泉這類充滿極端環境的區域中;然而之後人們發現古菌也生活在較不嚴酷的環境中,且現在認為比起細菌,古菌跟真核生物更為接近。
由於最後共同祖先的後代的基因庫、且所有的這些生物的DNA都遵循AT/GC法則且都使用同樣的二十種氨基酸之故,因此生物體間的水平基因轉移可行且常見。在1998年,卡爾·烏斯提出一項假說,這假說認為可能沒有任何生物可以被認為是最後共同祖先,而所有當代生物的基因遺傳是一群古代生物間的水平基因轉移造成的。
在2010年,由於「大量來自所有域的生物的分子序列現在變得可得之故」,因此研究人員出版了一項對共同祖先的正式測試結果,而這結果偏向支持所有生物有共同祖先的假說,而非上述的水平基因轉移假說等其他假說。基本生物化學原則使得所有生物源自單一的共同祖先的可能性,壓倒性地勝過其他的可能,而這是因為源自不同細胞生成事件的生物體發生水平基因轉移且不錯誤讀取彼此基因而不將之變成非編碼片段的狀況,是非常不可能發生的之故;此外,在目前發現已在蛋白質分子中發現的22種胺基酸之外,還有許多化學上可能用於蛋白質的胺基酸,而這與化學證據相合,將這點與形式統計測試相結合,可得出有單一的細胞生命是最後共同祖先的結論;另一方面,盡管研究壓倒性地支持所有生物源自單一的最後共同祖先的假說,這不表示這最後共同祖先是唯一的一種生物,相對地,最後共同祖先是多種早期微生物的其中一種,但卻是唯一一種在古太古代之後繼續存活的。
有證據顯示,細菌跟古菌在演化過程中其基因組發生簡化,而這表示說最後共同祖先可能比某些現代原核生物還複雜;貝葉斯系統發生比較認為最後共同祖先的基因組肯定是複雜的。
在F類三磷酸腺苷酶等極少數的情況中,相關基因的同線性可能比最後共同祖先還古老。
根所在的位置
目前廣泛接受的、根據多項分子生物學研究的生物演化樹的根位於細菌域這個單系群以及古菌及真核生物組成的演化支之間,而有很少數的研究將其根置於細菌域的厚壁菌門中,或認為綠彎菌門是古菌及真核生物組成的演化支以及其他種類細菌的基群(後面的假說由湯瑪斯·卡弗利爾-史密斯提出)。
馬丁在2016年的研究顯示地球上的生物源自類似海底熱泉的環境,但也有可能生命是在後期重轟炸期之類的事件後,才受限於如此環境的;對於最後共同祖先的基因的識別也受到批判,而這是因為這些基因可能是在之後才透過水平基因轉移在細菌跟古菌之間發生移轉的;此外另外,最後共同祖先有一氧化碳脫氫酶/乙醯輔酶A合酶的事實,除了跟這種生物可能是自營生物的假說相容外,也跟這種生物是混合營養的或異營生物的假說相容。
最後共同祖先的病毒
根據病毒在細菌與古菌這兩個主要的域當中的分布,最後共同祖先可能與已經相當複雜、且包括細菌與古菌的主要病毒種類的病毒組相關聯;此外,由於殼體蛋白的果凍捲結構可見於所有生物域的RNA與DNA病毒中之故,因此在最後共同祖先出現前病毒似乎就已發生大規模的演化;這些古代的病毒組可能以雙鏈去氧核糖核酸病毒域和多變去氧核糖核酸病毒域的雙鏈DNA病毒為主;此外,微小噬菌科病毒跟微管噬菌體目這兩類單鏈DNA病毒(也就是單鏈去氧核糖核酸病毒域的病毒)的起源可追溯到細菌的共同祖先;而古菌的共同祖先則最有可能受到梭狀病毒感染,而這幾類病毒存在於最後共同祖先病毒組但之後在其中一個域中消失的可能性是不能否認地;相反地,雖然從直觀想法來設想,最後共同祖先的病毒可能包含源自RNA世界的RNA病毒,但RNA病毒似乎並非最後共同祖先的一部分;而與之相對地,在最後共同祖先存活的時代,RNA病毒可能已很大程度地為更有效率的DNA病毒所取代。
參見
註解
外部連結
| |||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
