
| 演化生物学 |
|---|

|
人類演化过去一直指的是在旧的解剖学意义上的,根據演化學說,所做出的各種關於现代人出现的假设。通常包括灵长类的演化史。主要研究人属现代智人作为独特的人科物种出现的演化过程,而不是研究灵长类动物出现的演化过程。人类演化的研究涵盖多个学科领域,包括体质人类学、灵长类学、考古学、古生物学、行为学、语言学、进化心理学、胚胎学和遗传学。
在人類演化的脈絡下,「人類」這個專有名詞指的是「人屬」,但人類演化的研究往往包括其他人科動物,如南方古猿,人屬是在大約230萬至240萬年前的非洲,從南方猿人屬分支出來。科學家們估計,人類大約在距今500萬到700萬年之間,從他們與黑猩猩的共同祖先分支出來。其後發展的若干人屬物種和亞種,已告滅絕。這些包括棲息在亞洲的直立人、棲息在歐洲的尼安德塔人。古智人(早期智人)大約在距今25萬年到40萬年間演變出來。
關於解剖學上的現代人類的起源,科學家所持的主流觀點是「現代人類晚近非洲起源說」或稱「源出非洲」假說。此一演化說主張,智人興起於非洲,並且在大約距今5萬年到10萬年間遷移出非洲大陸,取代了在亞洲的直立人以及在歐洲的尼安德塔人。「源出非洲」的人類單一起源說,近期獲得粒線體DNA與Y染色體DNA兩項研究證據強力支持,成為國際主流普遍認同的人類演化起源說。
另一「現代人類多區域起源說」或稱「源起多地」假說的科學家主張,智人是在地理上相區隔但維持相互混血的諸多人類群體之間所產生的,這些群體源自於在距今約250萬年前,源出非洲的直立人所從事的一場遍及世界各地的遷徙。證據顯示,尼安德塔人的數個單倍群類型出現於目前在非洲以外的人類群體中;而且,尼安德塔人及其他人屬例如丹尼索瓦人可能貢獻了他們6%的基因體給現今的人類。
思想史
人屬的英文Homo這個詞,這個用來稱呼人類所屬的生物屬名,是拉丁文稱呼「人類」的字詞。它最初是由卡爾·林奈選來用在他的分類系統中。英文的「人類」一字human源自於拉丁文的humanus,這是Homo的形容詞。拉丁文的Homo源自於印歐語系的語根dhghem,意指「大地」。
卡爾·林奈及當時其他科學家也認為,大猿是人類最親密的近親,這是由於形態學和解剖上的相似處。人類與早期類人猿在譜系上具有連結的這種可能性,直到1859年查爾斯·達爾文出版《物種起源》之後,才可能變得更清楚。這本書主張支持新物種由早先物種演化而來的概念。達爾文的書沒有觸及人類演化的問題,只是說:「光芒將會投射在人類起源和他的歷史」。

最早關於人類演化本質的爭辯,出現在湯瑪斯·亨利·赫胥黎和理查·歐文之間。赫胥黎認為人類從類人猿演化而來,他列舉了人類和類人猿之間的許多相似性與相異性,特別是在他1863年的書《有關人類在自然界定位的證據》(Evidence as to Man's Place in Nature)。然而,許多達爾文的早期支持者(如阿弗雷德·羅素·華萊士和查理斯·萊爾)並不同意人類心智能力和道德情感的起源,可藉由自然選擇得到解釋。達爾文於1871年發表《人類的由來》一書時,將演化理論與性選擇運用於人類本身。
有一個嚴重的問題,就是缺乏介於人類與類人猿中間類型的化石。直到1920年代,這類化石才在非洲出土。1925年,雷蒙·達特(Raymond Dart)描述了南方古猿非洲種(Australopithecus africanus)的命名用標本(type specimen)是「陶格孩童」(Taung Child),這是在一座山洞裡發現的一具南方猿人嬰孩。這個孩童的遺骸包括一個保存完好的細小頭骨及其腦子的顱內腔模型(endocranial cast)。雖然這個腦子的腦容量較小(410立方公分),但是它的形狀是圓轉的,有別於黑猩猩和大猩猩的腦子,而更像是現代人類的腦子。此外,這個標本也顯示較短的犬齒,而且枕骨大孔的位置成為雙足直立行走的證據。所有這些特徵都使得達特相信,陶格孩童是一個雙足直立行走的人類祖先,也是介於類人猿與人類之間的一個過渡形式。
類人猿分類學(人類及其近親的分類)自從1950年代以來,已有很大的變化。例如,纖細種南方猿人曾被認為是現代人所屬的人屬(Homo)祖先。南方古猿與智人都是動物分類學上的人族(tribe Hominini)的一部分。
1970年代所蒐集的資料顯示,南方古猿是個多樣化的群體,南方古猿非洲種未必是現代人類的直接祖先。原先將南方猿人分類為「纖細種」與「粗壯種」,現在的重新分類將粗壯種放進單獨的屬:傍人。分類學家將人類、南方古猿及相關物種,以及其他類人猿放在同一個科,稱為人科(Hominidae)。
在1980年代與1990年代DNA測序的進展,特別是粒線體DNA以及稍後的Y染色體DNA的研究,增進了對人類起源的認識。來自世界各地廣大原住民群體的粒線體DNA與Y染色體DNA樣本排序結果,顯示了關於男性與女性基因遺傳的遠祖訊息。基因樹狀圖差異的校準,支持了現代人類晚近非洲起源說。分析呈現了在非洲各地具有最大的DNA模式多樣性,這顯示非洲是粒線體夏娃與Y染色體亞當的祖居地。
在人屬出現之前
大猿的演化

靈長類的演化史可追溯到距今6500萬年,牠們是在當時造成物種(特別是恐龍)滅絕的大災難下倖存的胎盤哺乳動物之中,最古老的一群。已知最古老的類靈長類哺乳動物物種,更猴來自北美,但牠們遍佈於歐亞大陸和非洲,在古新世和始新世期間,這兩處地方都處於熱帶。

隨著當代氣候的開始,特點是在大約距今3000萬年前的漸新世第一次出現了南極冰帽。在這個時間出現的靈長類動物是假熊猴。1980年代在德國發現的化石證據,並正在挑戰關於人類祖先源自非洲大陸的理論。
大衛·畢甘(David Begun)表示,這些靈長類動物在歐亞大陸蓬勃發展,而這個譜系導向非洲猿類和人類,包括森林古猿,從歐洲或西亞向南遷移至非洲。這支後來存活下來的熱帶群體,出土最完整的一群見於開羅西南部的法揚(Fayum)堆積層,始新世晚期和漸新世最早期的化石層位之中,這引發後續所有的現生靈長類,馬達加斯加的狐猴,東南亞的懶猴,非洲的嬰猴屬或「叢林嬰兒」(bush babies),以及人形亞目(Simian anthropoids);廣鼻猴次目(platyrrhine)或新世界猴,和狹鼻猴次目(catarrhine)或舊大陸猴、類人猿和人類。
已知最早的狹鼻猴次目,來自漸新世晚期的卡莫亞猿(Kamoyapithecus),在肯亞北部大裂谷的Eragaleit出土,定年為距今2400萬年前。其祖先一般認定是來自法揚的埃及猿、原上猿與傍猿(Parapithecus),在大約距今3500萬年前。在2010年,Saadanius 被描述為狹鼻猴次目冠狀群體(Crown group)最後共同祖先的一個近親,目前暫時定年到距今2900萬到2800萬年間,有助於填補長達1100萬年的化石記錄斷層。

在中新世早期,大約2200萬年前,來自東非的多種適應樹居生活的原始狹鼻猴亞目,顯示一段歷史悠久的早期多樣化發展。在2000萬年前的化石,包括歸類於最早的舊世界猴維多利亞猿(Victoriapithecus)的碎片。在這個屬當中,被認為處在猿類譜系之內,可推至距今1300萬年前的物種包括原康修爾猿、 Rangwapithecus、Dendropithecus、Limnopithecus、Nacholapithecus、Equatorius、Nyanzapithecus、非洲古猿、日猿(Heliopithecus)和肯尼亞古猿,都來自東非。其他中新世的普遍非cercopithecids來自遙遠的遺址,沉積在納米比亞洞穴的Otavipithecus,出自法國、西班牙和奧地利的加泰羅尼亞皮爾勞爾猿和森林古猿,證實在中新世早中期,在氣候相對溫暖的非洲和地中海盆地,有著廣大的生物型態多樣性。年代最近的中新世類人猿,山猿,出土自義大利距今900萬年的煤層。
分子證據指出,長臂猿(長臂猿科)的譜系,大約在距今1800萬至1200萬年前,從大猿的譜系分割出來;紅毛猩猩(猩猩亞科)的譜系,大約在距今1200萬年前,從大猿的譜系分割出來。目前並未出土足以清楚記載長臂猿祖先的化石,它可能源自於一個迄今未知的東南亞類人猿群體,但化石原紅毛猩猩可由來自印度的臘瑪古猿和來自土耳其的Griphopithecus做為其祖先的代表,追溯至距今1000萬年前。
人類譜系從其他大猿分裂兩族及後續演化
最接近大猩猩、黑猩猩和人類最後的共同祖先,可由肯亞出土的仲山納卡里猿和希臘出土的歐蘭猿為代表。分子證據顯示,在距今800萬至400萬年前,首先是大猩猩,然後是黑猩猩,從這條通往人類的演化路線分離出來;人類的DNA大約有98.4%與黑猩猩相同,這是比較單核苷酸多態性(single nucleotide polymorphisms)所得到的結果。大猩猩和黑猩猩的化石記錄相當有限。兩者的保存狀況都不理想(熱帶雨林土壤往往是酸性,會分解骨骼)。
其他人族有可能適應赤道帶以外的乾燥環境,還有羚羊、土狼、狗、豬、大象以及馬。約在距今800萬年前,赤道帶縮小。這些化石人族–從與黑猩猩分道揚鑣之後的人類譜系 - 比較廣為人知。最早的是乍得沙赫人(距今700萬年,乍得沙赫人可能是人與黑猩猩的最近共同祖先)和圖根原人(距今600萬年),其次是:
- 地猿(距今550萬到440萬年),包括地猿始祖種(Ardipithecus kadabba)和地猿根源種(Ardipithecus ramidus)。
- 南方古猿屬(距今400萬到180萬年),包括湖畔南方古猿、阿法南方古猿、南方古猿非洲種、羚羊河南方古猿、驚奇南方古猿,以及源泉南方古猿。
- 肯亞猿人屬(距今300萬到270萬年)。
- 傍人屬(距今300萬到120萬年),包括衣索比亞傍人、鮑氏傍人、粗壯種傍人(Paranthropus robustus)。
- 人屬(距今240萬年至今),包括巧人、盧多爾夫人、匠人、格魯及亞人、前人、西布蘭諾人、直立人、海德堡人、羅德西亞人、尼安德塔人、智人伊達圖亞種、古智人、弗洛瑞斯人。
人屬
智人是人屬唯一的現生物種,另有某些已告滅絕的人屬物種可能是智人的祖先,已經透過物種生成過程而分離出去(speciated),遠離我們祖先的路線。關於哪些群體應算作單獨的物種,哪些是亞種,目前尚未達成共識。在某些情況下,這是由於缺乏化石證據;在其他情況下,這是由於使用細微的差異,來分類人屬的各種不同物種的結果。「撒哈拉泵理论」(描述一個偶爾勉強稱得上「濕潤」的撒哈拉沙漠)提供對早期人屬變異的解釋。
依據考古學和古生物的證據,有可能在某種程度上推斷出,各種古代人屬的飲食習慣,並研究在人屬之中,飲食對於體質與行為演化所扮演的角色。
巧人
巧人存活在大約距今240萬到140萬年前。巧人是人屬的第一個物種,在上新世晚期或更新世早期的非洲南部與東非,大約距今250萬到200萬年前,從南方猿人分支出來。巧人具有比南方猿人更小的大臼齒與更大的腦袋,並且運用石頭(可能也用動物骨骼)製作工具。這是目前已知最早的人屬動物之一,其發現者路易·李基,因著牠與石器的關連性,暱稱為「巧人」。某些科學家提議將這個物種的屬別,從人屬移到南方猿人屬,因為依據對於牠的骨架所做的型態學分析,牠比較適應於樹居,而不是如同智人一般雙足直立行走。
卢多尔夫人與格鲁及亚人
這些物種名稱是用在大約從距今190到160萬年間的化石,然而牠們與巧人的關係尚未釐清。
匠人與直立人

第一個直立人化石是由荷蘭軍醫杜保斯(Eugene Dubois)在1891年於印度尼西亞的爪哇島發現的。他起初將這批化石材料命名為Pithecanthropus erectus,這是依據這些化石的型態學,他認為這是介於人類與大猿的中間物種。直立人存活的期間大約從距今180萬年到7萬年前(這或許指出,牠們可能受到距今約7萬年前的多峇巨災影響而滅絕;然而,直立人梭羅亞種(Homo erectus soloensis)與弗洛瑞斯人逃過這場巨災而存活下來。在前半段,從距今180萬到125萬年前的直立人,被認為是一個獨立的物種:匠人,或是直立人的亞種:直立人匠人亞種(Homo erectus ergaster)。
在更新世早期,距今150萬到100萬年間的非洲、亞洲與歐洲,某些巧人群體發展出較大的腦子,並製作更進步的石器;連同其他差異,足以讓人類學家將牠們分類為一個新物種:直立人。此外,直立人確實是第一個直立行走的人類祖先。這可能藉由膝蓋的演化,以及枕骨大孔(foramen magnum,人類頭骨正下方的大孔,連接腦子與脊髓)的位置前移而成為可能。直立人用火煮食肉類。
直立人的有名標本是北京猿人(因其出土化石數量龐大且層位較多);其他標本在亞洲(特別在印度尼西亞)、非洲與歐洲出土。現在有許多古人類學家用「匠人」來指稱這個群體的非亞洲型態,「直立人」這個名稱只保留用在亞洲所發現的化石,以呼應牠們在骨架與齒式方面有別於「匠人」之處。
西布蘭諾人與前人
這些物種可能是介於直立人與人屬海德堡種之間的中介物種。
海德堡人
海德堡人存活在大約80萬到30萬年前。學者也提議將其分類為智人海德堡亞種(Homo sapiens heidelbergensis)或智人古匈牙利亞種(Homo sapiens paleohungaricus)。
羅德西亞人與賈威斯頭骨
- 罗德西亚人,估計其年代約在距今30萬年到125,000年前。目前大多數的專家相信羅德西亞人是屬於人屬海德堡種,即使其他學者提議其為古智人與智人羅德西亞亞種(Homo sapiens rhodesiensis)。
- 在2006年2月,賈威斯頭骨(Gawis cranium)的化石出土,這可能是介於直立人與智人之間的中間物種,或是許多已告滅絕的演化路線之一。這個來自衣索比亞賈威斯的頭骨的年代約在距今50萬年到25萬年間。目前只有出版摘要資料,尚未由這個團隊發表經過同僚審查的研究報告。賈威斯頭骨的臉部特徵顯示,牠是一個中間物種或是「波多人」(Bodo man)女性的標本。
尼安德塔人

尼安德塔人存活的時間從距今40萬年前。也有學者提議將其分類為智人尼安德塔亞種(Homo sapiens neanderthalensis):學者持續爭辯「尼安德塔人」是否為一個單獨物種:人屬尼安德塔種,或者是智人的一個亞種。雖然這個爭論尚未平息,來自粒線體DNA排序的證據指出,在尼安德塔人與智人之間並未發生明顯的基因流佈。因此,這兩者是分別的物種,其共同祖先約在距今66萬年前。在1997年,馬克史東慶(Mark Stoneking)說道:「依據從尼安德塔人骨骼所萃取的粒線體DNA得到的結果指出,尼安德塔人並未貢獻粒線體DNA給現代人類…尼安德塔人並不是我們的祖先」。對於第二組尼安德塔人來源的調查,支持了這些發現。然而,在2010對於尼安德塔人基因體的排序指出,尼安德塔人確實在大約公元前75,000年與智人混血(在智人遷移離開非洲之後,但在他們分別遷往歐洲、中東與亞洲之前)。當代人類的DNA約有1%到4%源自尼安德塔人的DNA。這個1-4%的DNA差異只出現在非洲以外的人類。然而,「多區域演化」假說的支持者指出,最近研究顯示,非洲以外的核心DNA遺傳的時間可回溯到距今100萬年,即使這些研究的可靠性受到質疑。來自智人的競爭,可能導致尼安德塔人的滅絕。他們有可能在歐洲並存,直到距今1萬年前。
龙人
龙人化石1933年發現于現今的黑龍江省哈尔滨濱北大橋附近,由一队来自中国、澳大利亚和英国的学者, 通过研究这块哈尔滨古人类头骨化石,2021年6月25日在《The Innovation》杂志以封面文章形式发表了其研究成果,认定发现了新人种,命名为龙人。 龙人也是已灭绝的人类物种,该人种可能生活在大于14.6万年前,并且可能取代尼安德特人成为和现代人类亲缘关系最近的近亲。根据估测,智人、尼安德特人和龙人有可能早在100万年前就走向了不同的演化道路, 龙人和智人的共同祖先生活在大约95万年前。
智人
智人存活於距今25萬年到現在。在大約距今40萬年至25萬年間,當時是更新世第二間冰期,人類的腦容量變大,且發展出更細緻的石器製作技術,這些都是由直立人轉變為智人的證據。直接的證據顯示,有一股直立人從非洲向外遷移,接著在非洲的物種生成過程中,智人從直立人演化出來。智人後續在非洲內部與向非洲以外地區的遷移,最終取代了較早散居各地的直立人。這個遷移與起源理論往往稱為單地起源說或非洲起源說。目前的證據並未排除某些多區域演化或遷居的智人與各地既有的人屬群體混血的可能。這是目前古人类学的一個熱烈爭辯的領域。
當前的研究確認了,人類在遺傳學上是高度同質的;換言之,人類個體的DNA比起其他物種而言,具有較多的相似性。這可能源自人類演化的時間相對晚近,或由某些毀滅性的自然災害所引起的種群瓶頸,例如多峇巨災。然而,人類各群體已產生獨特的遺傳特徵,主要導因於人類的小群體遷移到許多新環境。這些因適應而產生的特徵,僅佔智人基因體的極小成分,但包括了各種不同特徵,例如膚色與鼻型,此外也有內部構造的特色,例如在高海拔地區更有效呼吸的能力。
來自衣索比亞的长者智人可能是一個已消失的亞種,存活於大約距今16萬年前。
弗洛瑞斯人
弗洛瑞斯人大約存活在距今10萬年到12,000年間,由於體型嬌小,被暱稱為哈比人,這可能是島內侏儒化所造成的結果。弗洛瑞斯人引人好奇的是牠們的體型與年代,這是一個具體的晚近的人屬物種,但展現了許多當代人類所沒有的衍生特徵(derived traits)。換言之,弗洛瑞斯人與當代人類具有共同祖先,但從當代人類的譜系分出去,依循一條獨特的演化路徑。主要的發現是一個年約30歲的女性骨架。在2003年出土,定年約在距今18,000年。這名女子存活時身高約一公尺,腦容量約380立方公分。(其腦容量如同黑猩猩,不到智人的平均值1400立方公分的三分之一。)
然而,學界持續爭辯著,究竟「弗洛瑞斯人」是否確實是個單獨存在的物種。目前有一些科學家相信,「弗洛瑞斯人」是受到病理侏儒症所苦的一種「智人」。這一假說受到部分的支持,因為在弗洛瑞斯島,也就是這個骨架被發現的島嶼,有某些當代人類是俾格米人。有人認為,這些較矮的俾格米人再加上病理侏儒症,確實可能創造出一種類似哈比人的人類。其他主要對於「弗洛瑞斯人」這個物種概念所做的攻擊,在於在其出土地點的工具僅僅與智人有關。
但是,這個病理侏儒症的假說無法解釋的是,為何弗洛瑞斯人其他的解剖特徵並不像現代人類(無論患病與否),反而很像我們種屬的古代成員。除了頭骨特徵之外,這些特徵也包括手腕、前臂、肩膀、膝蓋和腳的骨骼形式。
丹尼索瓦人
2008年,在西伯利亞阿爾泰山從事研究工作的考古學家,於丹尼索瓦洞穴發現一個小指骨頭碎片,來自目前稱為丹尼索瓦人群體的一位小女孩。在這處洞穴的相同層位出土的器物,包括一個手鐲,依據放射性碳定年法大約在距今四萬年前。由於丹尼索瓦洞穴氣候涼爽,因此在化石片段之中的DNA得以保存下來,目前已對其粒線體DNA及核心基因組DNA完成排序。
雖然其粒線體DNA的分歧點時間深度遠比預期更深,但是完整的基因組排序,顯示丹尼索瓦人與尼安德塔人屬於相同的世系,但兩者就在現代人類興起不久之後,就從同一條路線分裂開來。目前已知在歐洲,現代人類與尼安德塔人的重疊時間超過一萬年,而且這項發現引發一種可能性,尼安德塔人、現代人類、丹尼索瓦人或許曾經並存。帕博(Pääbo)指出,這個分支的存在,使得更新世晚期的圖像更加複雜。
人屬物種對照表
| 物種 | 生存年代(萬年前) | 生存地點 | 成人身高(米) | 成人體重(千克) | 腦容量(cm³) | 化石記錄 | 發現 / 名稱出版時間 |
|---|---|---|---|---|---|---|---|
| 人屬先驅種 Homo antecessor | 120 – 80 | 西班牙 | 1.75 | 90 | 1,000 | 2處遺址 | 1997 |
| 西布蘭諾人Homo cepranensis | 90 – 80? | 義大利 | 1,000 | 1個頭蓋骨 | 1994/2003 | ||
| 直立人Homo erectus | 150 – 20 | 非洲,歐亞大陸(爪哇,中國,印度,高加索) | 1.8 | 60 | 850(早期)– 1,100(晚期) | 多處遺址 | 1891/1892 |
| 匠人Homo ergaster | 190 – 140 | 東非與南非 | 1.9 | 700–850 | 多處遺址 | 1975 | |
| 弗洛瑞斯人Homo floresiensis | 10? – 1.2 | 印尼 | 1.0 | 25 | 400 | 7具個體 | 2003/2004 |
| 豪登人Homo gautengensis | >200 – 60 | 南非 | 1.0 | 2010 | |||
| 人屬喬治亞種 Homo georgicus | 180 | 格鲁吉亚 | 600 | 4具個體 | 1999/2002 | ||
| 巧人Homo habilis | 230 – 140 | 非洲 | 1.0–1.5 | 33–55 | 510–660 | 多處遺址 | 1960/1964 |
| 人屬海德堡種 Homo heidelbergensis | 60 – 35 | 歐洲,非洲,中國 | 1.8 | 60 | 1,100–1,400 | 多處遺址 | 1908 |
| 尼安德塔人Neanderthal | 35 – 3 | 歐洲,西亞 | 1.6 | 55–70 | 1,200–1,900 | 多處遺址 | (1829)/1864 |
| 人屬羅德斯種 Homo rhodesiensis | 30 – 12 | 尚比亞 | 1,300 | 非常少 | 1921 | ||
| 人屬魯道夫種 Homo rudolfensis | 190 | 肯亞 | 1個頭骨 | 1972/1986 | |||
| 智人伊達圖亞種 Homo sapiens idaltu | 16 – 15 | 衣索比亞 | 1,450 | 3個頭蓋骨 | 1997/2003 | ||
| 現代人類Homo sapiens sapiens | 20 – 現代 | 世界各地 | 1.4–1.9 | 50–100 | 1,000–1,850 | 现存 | —/1758 |
使用工具




使用工具已被學者詮釋成一種智力的標誌,而且學者提出理論,工具使用可能觸發了人類演化的某些層面----最值得一提的是人類腦子的持續擴張。古生物學目前依然未能解釋腦子經過數百萬年的擴張,即使說從能量消耗來看,腦子的需求非常大。當代人類的腦子消耗約20瓦(每天400千卡),佔人體能量消耗的五分之一。工具使用的增加,使得人類能獵得能量豐富的肉類,並有能力處理更多能量豐富的植物產品。研究人員認為,早期人類是在演化壓力下,提高他們創造和使用工具的能力。
值得一提的是,有許多物種能夠製作和使用工具,但人類這個物種控制了製作的範圍並使用更複雜的工具。目前已知最古老的工具來自衣索比亞的「奧度瓦伊石器」(發現地點為奥杜威峡谷)。這些工具的年代距今250萬年到260萬年,來自已知最早的「人屬」物種。目前尚無證據顯示,距今250萬年前有任何「人屬」標本。在一些奧度瓦伊工具附近,出土了一個人屬化石,值得注意的是其定年在距今230萬年,這隱約顯示,也許早期人屬物種確實創造和使用這些工具。這確實可能,但沒有堅實的證據。伯納伍德(Bernard Wood)指出,「傍人屬」與這個「奧度瓦伊技術叢」("Oldowan Industrial Complex")大致同時存在這個區域。雖然沒有直接證據可佐證傍人屬是工具製造者,但他們的解剖構造提供了間接證據,顯示他們具備這方面的能力。大多數古人類學家都同意,確實最有可能是早期的「人屬」物種製作出奧度瓦伊工具。他們認為,當大多數的奧度瓦伊工具與人類化石同時被發現時,出現的始終是人屬,但不是傍人屬。
1994年,蘭德爾·蘇斯曼採用人屬及傍人屬的對向性拇指來做為他的立論基礎,推論他們都是工具製造者。他比較了人類和黑猩猩的拇指骨骼和肌肉,發現人類具有三條黑猩猩所欠缺的肌肉。人類也有較厚實的掌骨與更大的頭部,這使得人類的手在精準抓握方面,比黑猩猩更成功。蘇斯曼提出辯護,人類拇指的現代解剖學構造是基於製作與操作工具的需求而產生的演化回應,這兩個物種確實是工具製造者。
石器
經證實,最早的石器約在距今260萬年前,當時在東非的巧人使用的工具,是由圓形礫石簡單捶打而成的礫石器、砍器。這標示了舊石器時代的開始,結束在最後一次冰河時代告終之時,大約距今一萬年前。舊石器時代細分為舊石器時代早期(約在距今35萬至30萬年前結束),舊石器時代中期(直到距今五萬年到三萬年前)和舊石器時代晚期。
從距今70萬至30萬年的這段時期,稱為阿舍利傳統,當時的匠人(或直立人)由燧石和石英石製作手斧,起初相當粗糙(早期阿歇爾),後來經過在石片兩側更多細部修整。在距今35萬年之後,更細緻的勒瓦娄哇技法發展出來。它包含一系列連續敲擊,製作出刮削器、切割器、針、扁針。最後,大約在距今五萬年之後,尼安德塔人及克羅馬儂人製作出更細膩與專門化的燧石工具(刀、石瓣器)。在此期間,他們開始製作骨製工具。
朝向行為現代性的轉型
直到大約距今5萬到4萬年前,石器的使用似乎逐步取得進展。每個階段(巧人、匠人、尼安德塔人)從比起前一個階段更高的層次開始,但每個階段開始後,進一步發展就遲緩下來。當前的古人類學家,正在爭辯這些智人物種究竟是否具備跟現代人有關的某些或許多文化和行為特徵,諸如語言、複雜的象徵思維、科技創造力等等。看起來,他們有很長一段時間,是維持簡單的技術和覓食模式的文化保守分子。
大約距今5萬年前,當代人類文化開始更快速地演進。這項朝向行為現代性的轉變,稱為歐亞大陸人的「大躍進」,或稱為「舊石器晚期革命」,這是由於在考古學記錄中,突然大量出現當代行為的跡象。某些學者認為這個轉型是更為漸進的,花費更長時間,因為有些特徵早在距今20萬年前的古非洲智人當中。
現代人類開始埋葬死者、製作衣服、發展複雜的狩獵技巧(如使用誘捕坑或逼使動物跳下峭壁),並從事石洞壁畫。隨著人類文化的進展,人類的不同人群推出新穎的技術,加入既有的技術當中:魚鉤、鈕扣與骨針等等的器物,呈現了不同人類群體之間的變異,某些東西未曾出現在距今5萬年前的人類文化當中。一般來說,尼安德塔人的各個群體在技術方面並未有所差異。
人類學家指出的現代人類行為的具體例證,包括專業化的工具、珠寶首飾和圖像的使用(如洞穴壁畫)、生存空間的組織、儀式(例如,有陪葬品的墓葬)、專門的狩獵技術、探索較不適合人居的地理區域、以物易物的貿易網絡等等。學者之間的爭辯還在持續進行,圍繞在究竟是一場「革命」導致現代人類的產生,或者說這個演變是漸進的。
人類演化的模型
今天,所有的人類都同屬單一的群體:智人智人亞種(Homo sapiens sapiens),並沒有物種區隔。然而,根據「源出非洲」模型,這並不是人屬的第一個物種:人屬的第一個物種「巧人」,在至少距今兩百萬年演變出來,巧人生活在非洲不同區域的時間相對較短,「直立人」在超過距今180萬年前演變出來,到了距今150萬年前就已散布到舊世界各地。
人類學家對於當代人類群體演化有著分歧的看法,是否只在東非發生演變、物種形成,然後遷移到非洲以外,並取代歐亞大陸的人類群體(所謂的「源出非洲」模型或「完全替代」模型);或是演變成一個相互連結的群體(由「多區域演化」假設所假定的模型)。
源出非洲
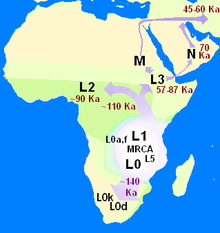
依據由克里斯·斯特林格和彼得·安德魯斯所發展的源出非洲模型(Out-of-Africa model),現代智人是在距今20萬年前的非洲演變而來的。智人大約在距今七萬年到五萬年間,開始從非洲遷移出來,並最終取代當時存在於歐洲和亞洲的原始人屬物種。源出非洲已獲得研究的支持,包括對女性的粒線體DNA和男性Y染色體的研究。使用133個類型的粒線體DNA構建譜系樹狀分析後,研究人員得出結論,她們都是來自非洲的同一個女人的後代,這個女人稱為粒線體夏娃。源出非洲模型也從非洲人群具有最高的粒線體遺傳多樣性這項事實得到支持。
關於這個向外遷徙的過程究竟是一次或多次,學者抱持著不同的理論。一套多次遷徙模型,其中包括向南擴散理論,近年來已經獲得來自基因、語言和考古證據的支持。在這個理論中,大約在距今七萬年前,現代人類從非洲之角展開一場沿著海岸的遷徙。這群人有助於將人類擴展到東南亞和大洋洲,這解釋了為何早期人類在這些區域的遺址年代,遠遠早於在黎凡特的遺址年代。人類的第二波遷徙浪潮跨越西奈半島進入亞洲,產生了在歐亞大陸的大批人群。第二波的人群擁有更成熟的工具技術,而且比起第一波的人群更少依賴沿海的食物來源。第一波人群的擴張證據可能已被每一次冰河時期結束時,上升的海平面所摧毀。這個多次遷徙模型與來自粒線體DNA的研究結果相衝突,後者顯示歐亞大陸的人群以及東南亞與大洋洲的人群都是從同一個粒線體DNA譜系傳承下來,這支持了單一的源出非洲遷徙過程,產生了所有在非洲以外的人類群體。
莎拉·提胥可夫率領的廣大非洲的遺傳多樣性研究團隊,從113個不同人群的採樣當中,發現布希曼人具有最大的遺傳多樣性,這使得他們成為14個人類基因叢集的其中一個。這項研究也定位了非洲西南部現代人類遷移的源頭,接近納米比亞和安哥拉海岸。
根據人類學家和考古學家所採用的多峇巨災理論,大約在距今七萬年前,印尼蘇門答臘島的多峇湖發生一場超級火山噴發,造成了全球性的後果,致使當時的大多數人死亡,並創造出一個種群瓶頸,影響現今所有人類的基因遺傳。
多地起源模型
「多地起源模型」是一種「解釋人類演化的模型」,由沃波夫(Milford H. Wolpoff)於1988年提出。多區域演化的觀點主張,人類演化是從距今250萬年前的更新世開始,直到現在,一直維持著單一的、連續的人類物種,在世界各地演化成為現代智人。依據多區域假設,化石及基因體資料是說明全球性人類演化的證據,這與最近由晚近非洲起源說所假定的物種形成過程相矛盾。理查德·李奇認為化石證據並不充足,無法解決這個爭論。 對於Y染色體及粒線體DNA的單倍群研究已大大支持一個晚近的非洲起源。來自常染色體DNA的證據也支持晚近非洲起源的可能性。然而,某些研究顯示,古智人(Archaic Homo sapiens)的遺傳成分也有可能摻雜在當代人類當中。最近關於尼安德塔人與丹尼索瓦人的基因體排序也顯示確實發生某些混雜。在非洲以外的現代人類在其基因體中,有2-4%的尼安德塔人的等位基因,美拉尼西亞人額外具有4-6%的丹尼索瓦人等位基因。這些新的結果並不與《源出非洲》模型相抵觸,但顯示發生了一些額外的混雜事件。可能源自於距今73,000年前的多峇超級火山災難的一個種群瓶頸之後,經過一段復甦時間,一支很小的群體離開非洲,並遇到尼安德塔人,可能是在中東,他們的後代擴散到世界各地,有一小支遇到了丹尼索瓦人,可能是在東南亞,就在他們遷入美拉尼西亞之前。
外部链接
- 紀錄片-人類起源之謎 (页面存档备份,存于互联网档案馆)
- http://sa.ylib.com/MagCont.aspx?Unit=featurearticles&id=1499(页面存档备份,存于互联网档案馆)
- (简体中文)songshuhui.net网站上关于人類起源的介绍 (页面存档备份,存于互联网档案馆)
- (英文)BBC: The Evolution of Man (页面存档备份,存于互联网档案馆)
- (英文)BBC: Finds test human origins theory (页面存档备份,存于互联网档案馆)
- (英文)Researchers say human foot not unique, more like those of great apes (页面存档备份,存于互联网档案馆)
参见
| ||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


| ||||||||||||||||||||||||||||||||